Конфигурация - цепь
Cтраница 2
Влияние ограничений конфигурации цепи на процессы кристаллизации и плавления зависит от статистической суммы ( при условии, разумеется, что ее критическое значение, определенное выше, находится в доступной температурной области); для реальных полимеров требуется вычисление со с подстановкой потенциалов заторможенного вращения. [16]
Для доказательства предложенной конфигурации складчатой цепи Мэнли искал меридианные рефлексы малоуглового рентгеновского рассеяния, соответствующие периодам около 35 А. [17]
 |
Модель цепи ПТФЭ. [18] |
Определяющую роль в конфигурации цепи в кристаллическом ПТФЭ играют внутримолекулярные, а не межмолекулярные силы взаимодействия [60], спиральная структура упаковы. [19]
 |
Модели полимеров в растворе. [20] |
Очевидно, что конфигурация цепи полностью определяется химической историей полимера и не зависит от физического состояния. Конформация же, напротив, определяется в основном рядом физических факторов. [21]
Необходимую информацию относительно конфигурации полиеновой цепи удается получить с помощью изучения спектров люми-нисценции образцов полипропиоловой кислоты. С помощью этого метода было доказано, что полимеризация пропиоловой кислоты в твердой фазе приводит к образованию полимера преимущественно гранс-трансоидной конфигурации основной цепи сопряжения. В случае жидкофазной полимеризации образуются макромолекулы грамс-цисоидной или цмс-трансоидной конфигурации. [22]
Наличие внутренних флуктуации конфигурации цепей должно приводить к уменьшению силового поля между частицами, а также к изменению геометрии частиц. Ли и др. [104] недавно опубликовали теоретический анализ, который качественно описывает основные результаты динамического рассеяния света в растворах ДНК. Использованная ими гидродинамическая модель основана на предположении, что мгновенная форма молекул ДНК несферична [105] и что, следовательно, в концентрированных растворах имеется сильная анизотропия трансляционной диффузии. Когда q - 0, рассеяние целиком обусловлено медленной модой, постоянная времени которой определяется средним коэффициентом диффузии. Такой анализ предполагает, что внутренняя мода релаксации не зависит от межмолекулярного взаимодействия и что внутричастичный интерференционный вклад в интенсивность рассеяния пренебрежимо мал. Авторы также показали, что не существует такого гидродинамического механизма, по которому независимо ориентированные молекулы могли бы вызвать релаксационную компоненту более медленную, чем трансляционная диффузия. Далее они показали, что взаимодействие, характерное для твердых сфер и соответствующее дальнему порядку, не может быть источником аномальной рассеивающей компоненты. [23]
А-константа, определяемая конфигурацией цепи в данном растворителе. Разумеется, формула ( 33) верна лишь в первом приближении ( по достижении М величины ЫЛА величина t не падает внезапно до нуля), однако изложенные соображения довольно наглядно показывают возможный путь интерпретации МБР при полимеризации на комплексных металлорганических катализаторах. [24]
Аналогичным путем были определены конфигурации цепей i а-г ттаперче и в натуральном каучуке. [25]
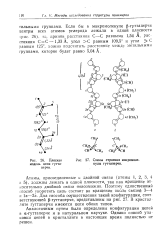 |
Схемы строения макромолекулы гуттаперчи. [26] |
Аналогичным путем были определены конфигурации цепей в а-гуттаперче и в натуральном каучуке. Однако способ упаковки цепей в кристаллите в настоящее время полностью не решен. [27]
В результате имеет место совершенно беспорядочная конфигурация цепи. [28]
Нужно подчеркнуть, что сильно свернутые конфигурации цепей, где объемные эффекты особенно существенны, автоматически исключаются в сеточной теории благодаря наличию фиксированных точек, положение которых определяется внешними размерами образца. [29]
Аналогичным путем были определены конфигураций цепей в а-г ттаперче и в натуральном каучуке. Однако способ упаковки цепей в кристаллите в настоящее время полностью не выяснен. [30]
Страницы: 1 2 3 4