Конфигурация - боковая цепь
Cтраница 1
Конфигурация боковой цепи при С17 точно не установлена, но принято считать, что последняя в природных стероидах занимает р-положение. Пространственное расположение боковой цепи, однако, имеет решающее значение, так как соединения с 17 р-цепью активнее их 17а - эпимеров. Ниже описаны наиболее важные сердечные гликозиды. [1]
Если наблюдаемое отклонение не связано с неоднородностью образца, то наиболее вероятно, что оно обусловлено идентичностью конфигурации боковых цепей, поскольку энантиомеры, как известно, должны давать точно асимметрические кривые. [2]
При осуществлении целей, поставленных в приведенных работах, решались задачи синтеза оксициклопенте-нонов заданной структуры ( 4 - или 5-оксизамещенные Д2 - циклопентеноны) и стерической ( цис -, транс -) конфигурации боковых цепей. При этом был разработан ряд эффективных методов, ставших достоянием синтетической химии. Сюда относятся, например, методы получения ранее недоступных 4-оксициклопентенонов. [3]
При lla - гидроксилировании 16-дегидропрогестерона ( LI) культурами Aspergillus niger [104] или Rhizopus nigricans [105] имеет место также восстановление А1в - связи с образованием соединения ( LII), имеющего 17а - конфигурацию боковой цепи. [4]
Это вещество, подобно биотину, содержит три асимметрических атома углерода и должно существовать в восьми оптически активных и четырех рацемических модификациях; в двух рацематах положение аминогруппы должно соответствовать цис -, а в двух остальных транс-расположению. Конфигурация боковой цепи, связанной со вторам углеродным атомом цикла, не была установлена. [5]
Ввиду недостаточной точности спектральных данных при оценке относительных вкладов различных донорных лигандов соответствие сдвига энергии с координационным окружением в настоящее время не может быть проанализировано на количественном уровне. Сравнение соответствующих геометрических свойств металламино-кислотных комплексов в табл. 4 подтверждает, что геометрия координации и конфигурация боковых цепей координированных аминокислот, вероятно, либо различаются очень незначительно, либо совершенно одинаковы. [6]
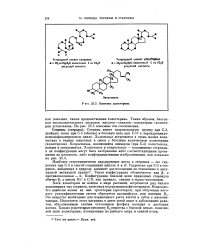 |
Биогенез холестерина. [7] |
Наиболее стереохимически варьирующие места в стеринах - это гид-роксил при С-3 и способ соединения циклов А и В. Такая конфигурация обозначается как р а противоположная - а. Конфигурация боковой цепи природных стероидов обычно р, а циклы В С и CJD, как правило, спаяны в траке-положении. [8]
Наиболее стереохимически варьирующие места в стеринах - это гид-роксил при С-3 и способ соединения циклов А и В. Такая конфигурация обозначается как ( 3, а противоположная - а. Конфигурация боковой цепи природных стероидов обычно р, а циклы BJC и CjD, как правило, спаяны в траке-положении. [9]
Ошибки при использовании изоморфных производных, полученных путем замены иона металла, находящегося в нативном белке, на более тяжелые элементы, могут возникать из-за изменений в структуре металл-лигандных центров при замене металла. Кристаллографические исследования карбоксипептидазы А [29, 67], карбоангидразы С [68, 69] и нуклеазы стафилококка [33] показали, что более тяжелый ион металла смещен относительно металла в нативном белке, как и следовало ожидать, исходя из различных координационных свойств и ионных радиусов соответствующих металлов. Из-за вполне вероятного в этих условиях изменения конфигурации боковых цепей аминокислот, являющихся лигандами, отсутствие точного изоморфизма приводит к некоторой ошибке в расчете фаз. Эти изменения также могут привести к утрате точного изоморфизма. Вообще говоря, эта проблема может быть частично решена путем использования фаз, полученных для нескольких изоморфных производных. При расчете электронной плотности вблизи центра связывания металла для производного, в котором тяжелый атом замещает катион нативного белка, могут быть использованы фазы, определенные для других производных. [10]
Страницы: 1