Ассимиляция - углекислота
Cтраница 2
У всех исследованных аэробных и факультативно анаэробных хе-молитотрофов ассимиляция углекислоты в автотрофных условиях происходит в результате действия рибулезодифосфатного цикла, называемого также циклом Кальвина. [16]
Освобождающаяся при этом окислении энергия используется ими для ассимиляции углекислоты. Железобактерии в очистных сооружениях встречаются в исключительных случаях. [17]
Растения синтезируют жиры из крахмала, который является продуктом ассимиляции углекислоты. [18]
Растения синтезируют жиры из крахмала, который является продуктом ассимиляции углекислоты. В животном организме жиры образуются из жиров пищи, а также из углеводов. [19]
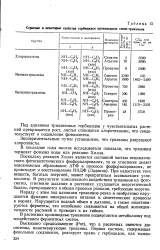 |
Строение и некоторые свойства гербицидов производных симм-триазинов. [20] |
При недостатке этих веществ, богатых энергией, может прекратиться ассимиляция углекислоты. В результате комплексного воздействия триазинов на фотосинтез, угнетение дыхания в растениях ухудшается энергетический баланс, прекращаются в общем процессе реакции, требующие энергии. [21]
Другие примеры взаимных превращений Сахаров уже рассматривались при изложении процессов ассимиляции углекислоты в фотосинтетическом цикле, с некоторыми из них дополнительно познакомимся при изучении биохимии дыхания и окислительного распада углеводов. Все сказанное показывает, что процессы обмена углеводов очень тесно взаимосвязаны. [22]
В условиях интенсивного роста фототрофных бактерий большая часть первичных продуктов ассимиляции углекислоты и органических соединений используется на синтез белков и других важнейших компонентов клеток. Но при задержке роста культур в результате недостатка фосфора и особенно азота происходит образование в больших количествах запасных веществ. И пурпурные и зеленые бактерии могут накапливать в качестве запасных продуктов полисахариды типа гликогена. Накопление того или другого запасного соединения зависит не только от особенностей организма, но и от источника углерода. Полисахариды преимущественно накапливаются пурпурными бактериями при культивировании их на минеральных средах, а также в присутствии пирувата, пропионата, малата или сукцината. При использовании бутирата или ацетата, особенно в отсутствие бикарбоната, происходит накопление почти исключительно поли-р-оксибутирата, так как синтез его начинается с конденсации двух молекул ацетил - КоА с образованием аце-тоацетил - КоА и далее р-оксибутирата. [23]
Часть образуемой при этом энергии доступна для использования в процессе ассимиляции углекислоты. [24]
При еще более низких концентрациях сернистый ангидрид не влиял на ассимиляцию углекислоты, за исключением тех случаев, когда окуривание продолжалось слишком долго. [25]
В настоящей статье я изложу новую гипотезу, объясняющую химический механизм ассимиляции углекислоты проще и правдоподобнее, чем изложенные выше гипотезы, и приводу опыты, которые я поставил для ее проверки. [26]
Из данных, приведенных в табл. 9, видно, что процесс ассимиляций углекислоты протекал интенсивнее на вырубке, чем под пологом леса. [27]
Изложенные соображения привели меня к следующей гипотезе, хорошо объясняющей химический механизм ассимиляции углекислоты растениями и подтверждающейся рядом экспериментальных данных. [28]
Отметим также, что у зеленых нитчатых бактерий Chloroflexus было показано, что ассимиляция углекислоты осуществляется не только в цикле Кальвина, но и необычными путями - в 3-гидрокси-пропионатном цикле и / или восстановительном цикле дикарбо-новых кислот. [29]
Как видно, высказанные до сих пор гипотезы, пытающиеся объяснить химический механизм ассимиляции углекислоты, лишь отодвинули вопрос - проблема же, как и раньше, остается неразрешенной. Большая заслуга этих гипотез заключается, однако, в том, что вопрос правильно поставлен в плоскости химических реакций; это единственный путь, который может привести к его удовлетворительному решению. [30]
Страницы: 1 2 3 4