Двойной кроссинговер
Cтраница 1
Двойной кроссинговер ( Double crossingover) Кроссинго-вер, происходящий одновременно в двух точках пары гомологичных хромосом. [1]
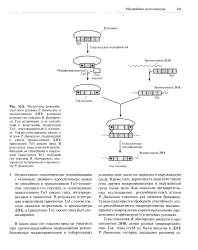
Осуществили гомологичную рекомбинацию с помощью двойного кроссинговера между не способным к транспозиции Тп5 - элемен-том, несущим ген токсина, и плазмидным транспозоном Тп5 дикого типа, интегрированным в хромосому. В результате интеграции измененный транспозон Тп5 с геном токсина оказался встроенным в хромосомную ДНК, а транспозон Тп5 дикого типа был элиминирован. [3]
 |
Генная ( или делеций. [4] |
Продукт генной конверсии формально аналогичен результату двойного кроссинговера, но отличается отсутствием второе, реципрокного, продукта рекомбинации. [5]
Если клетка трансформирована нереплицирующейся плазмидой, несущей клонированный ген в середине клонированного фрагмента с хромосомным сайтом интеграции, то может произойти спаривание между гомологичными нуклеотидными последовательностями плазмиды и хозяйской ДНК ( рис. 6.15, А) и далее интеграция в результате двойного кроссинговера, осуществляемого ферментами клетки-хозяина. [6]

Культуру клеток насекомого, трансфициро-ванную ДНК AcMNPV, трансфицируют затем транспортным вектором, несущим клонированный ген. В некоторых дважды трансфицирован-ных клетках происходит двойной кроссинговер, в результате которого клонированный ген вместе с промотором и сигналом терминации транскрипции гена полиэдрина встраивается в ДНК AcMNPV ( рис. 7.9), замещая ген полиэдрина. Вирионы, не содержащие этого гена, образуют зоны клеточного лизиса, из которых можно выделить рекомбинантный бакуловирус. [8]

В результате двойного кроссинговера между вектором и геном AcMNPV клонированный ген встраивался в последний и попадал под контроль сильного промотора, функционирующего на последних стадиях литического цикла. Клетки насекомого, инфицированные рекомбинант-ным бакуловирусом, синтезировали гетероло-гичный белок. [10]
В результате двойного кроссинговера в некоторых клетках восстанавливался кольцевой геном AcMNPV, который содержал клонированный ген и функциональный ген, необходимый для осуществления литического цикла. [11]

Исходная методика получения рекомбинантных бакуловирусов в дальнейшем была изменена по ряду причин. Во-первых, применение промотора гена полиэдрина имеет ограничения: белки, синтезирующиеся на поздней стадии литиче-ского цикла, часто оказываются модифицированными не до конца. Для решения этой проблемы промотор гена полиэдрина заменили одним из сильных промоторов AcMNPV, активно функционирующих с самого начала и до конца литического цикла. Во-вторых, линеаризация генома AcMNPV перед трансфекцией клеток насекомого увеличивает долю зон лизиса с реком-бинантными вирусами. Расщепление генома AcMNPV в одном сайте уменьшает число зон с нерекомбинантными вирусами, потому что линеаризованные геномы бакуловирусов обладают ограниченной инфицирующей способностью. В результате двойного кроссинговера между линеаризованной ДНК AcMNPV и кольцевым транспортным вектором образуется замкнутая кольцевая молекула, которая обладает инфицирующей способностью. [13]
Страницы: 1