Афферент
Cтраница 1
 |
Мышечное веретено. [1] |
Афференты группы II полисинап-тически возбуждают мотонейроны сгибателей и тормозят мотонейроны разгибателей. Имеются, однако, данные, что афферснты в группе II, идущие от разгибателей, могут возбуждать мотонейроны своей мышцы. [2]
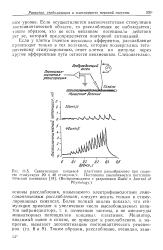
Если у клетки имеется несколько афферентов, расслабление происходит только в тех волокнах, которые подвергались тета-ническому стимулированию, ответ клетки на импульс через другие афферентные пути остается неизменным. [4]
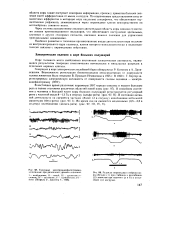
По современным представлениям вход от мышечных афферентов в моторную кору настолько специфичен, что обеспечивает осуществление рефлексов, замыкающихся через пирамидные клетки непосредственно на мотонейронах спинного мозга. [6]
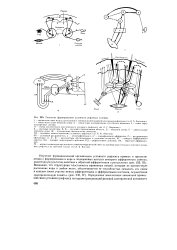 |
Гипотезы формирования условного рефлекса ( схемы. [7] |
УР - условный раздражитель; а - неспецифические и а2 - специфические афференты; О - раздражители обстановки; А, А1, А2 - анализаторы обстановочных и пусковых сигналов; АС - аппарат афферентного синтеза; АД - аппарат акцепторного действия; РД - рефлекторное действие; ОА - обратная афферента-ция. [8]
И без растяжения мышцы, активация у-эфферентов сама по себе вызывает им-пульсацию афферентов веретен вследствие сокращения интрафузальных мышечных волокон. [9]
 |
Послойная топография серого вещества спинного мозга по Рекседу. [10] |
В целом нервные клетки первой - четвертой пластин захватывают всю вершину дорсального рога и образуют первичную сенсорную область спинного мозга. Сюда проецируются волокна большей части дорсально-корещковых афферентов от экстерорецепторов, включая кожную и болевую чувствительность. В этой же зоне локализованы нервные клетки, дающие начало нескольким восходящим трактам. [11]
При анализе установленной в наших опытах устойчивости ранних компонентов ассоциативных ответов к яду кобры и фармакологическим агентам необходимо учитывать особенности морфофункциональной организации теменной ассоциативной области по сравнению с проекционными. Известно, что ассоциативная кора отличается от проекционной отсутствием преобладания специфических афферентов в каком-либо одном уровне ее поперечника, более диффузным распределением их по вертикали и горизонтали, а главное значительно меньшим числом рассматриваемых афферентов ( О. С. Андрианов, А. В противоположность этому волокна, идущие из тех же специфических ядер-реле таламуса в проекционные области, направляются мощным концентрированным пучком к слою IV и нижнему подслою слоя III. Отсюда следует, что ранние компоненты ассоциативных ответов формируются более скудным и рассеяным потоком специфической афферентации по сравнению с тем, который генерирует первичный ответ. Указанные мор-фофункциональныс особенности ассоциативной коры, очевидно, в большей степени определяют более высокий порог чувствительности ранних компонентов ассоциативных ответов к нейротропным агентам. [12]
При анализе установленной в наших опытах устойчивости ранних компонентов ассоциативных ответов к яду кобры и фармакологическим агентам необходимо учитывать особенности морфофункциональной организации теменной ассоциативной области по сравнению с проекционными. Известно, что ассоциативная кора отличается от проекционной отсутствием преобладания специфических афферентов в каком-либо одном уровне ее поперечника, более диффузным распределением их по вертикали и горизонтали, а главное значительно меньшим числом рассматриваемых афферентов ( О. С. Андрианов, А. В противоположность этому волокна, идущие из тех же специфических ядер-реле таламуса в проекционные области, направляются мощным концентрированным пучком к слою IV и нижнему подслою слоя III. Отсюда следует, что ранние компоненты ассоциативных ответов формируются более скудным и рассеяным потоком специфической афферентации по сравнению с тем, который генерирует первичный ответ. Указанные мор-фофункциональныс особенности ассоциативной коры, очевидно, в большей степени определяют более высокий порог чувствительности ранних компонентов ассоциативных ответов к нейротропным агентам. [13]
Шейбел и Шейбел ( 1967, рис. XV-3 и XV-4) также описали эту систему нейронов; по мнению этих авторов, она функционирует как система неспецифических афферентных путей к коре больших полушарий. Характерно, что аксоны этих клеток длинные и на их окончаниях имеются разветвления, обвивающие апикальные дендриты пирамидных клеток, свободно соединяющиеся с дендритами, в противоположность хорошо ограниченным синапсам, типичным для специфических афферентов. [14]
Сенсорная слуховая область отличается богатством клеточного и волоконного состава, имеет хорошо выраженный слой крупных пирамидных нейронов. Типичным для слуховой коры является также хорошо выраженный слой звездчатых клеток. В слуховую область, кроме слуховых путей, проецируются также вестибулярные афференты. [15]
Страницы: 1 2