Лигнин - срединная пластинка
Cтраница 1
Лигнин срединных пластинок и лигнин вторичных слоев извлекаются из растительных материалов не одинаково легко. Часть лигнина химически связана с углеводами, и для ее отделения от них необходимо химическое воздействие на материал. [1]
 |
Растворение лигнина. [2] |
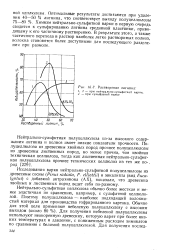
Химизм нейтрально-сульфитной варки в первую очередь сводится к сульфированию лигнина срединной пластинки, приводящему к его частичному растворению. В результате этого, а также частичного перехода в раствор наиболее легко растворимых полиоз, волокна становятся более доступными для последующего разделения при размоле. [3]
Оказалось, что в лигнине вторичных стенок волокон и лучевых клеток древесины березы ( Betula papyrifera) наблюдается высокое относительное содержание сирингильных единиц, а лигнины сложной срединной пластинки и стенок сосудов богаты гваяцильными единицами. В еловых ( Picea abies) трахеидах лигнин вторичной стенки содержит в 2 раза больше фенольных ОН-групп по сравнению с лигнином сложной срединной пластинки. [4]
Исходные монолигнолы включаются в протолигнин в том же порядке, в каком они образуются в ходе биосинтеза - сначала - кумаровый, затем конфериловый спирт и, наконец, синаповый. Поэтому гваяцильный лигнин хвойных неоднороден: лигнин срединной пластинки содержит большую долю Н - единиц, чем лигнин вторичной стенки, тогда как S-еди-ницы обнаруживаются в лигнине вторичной стенки ближе к полости. Состав гваяцил-сирингильного лигнина лиственных подчиняется этой же закономерности, но, соответственно, содержит меньшую долю Н - единиц ( только в лигнине срединной пластинки) и большую долю S-единиц. [5]
Оказалось, что в лигнине вторичных стенок волокон и лучевых клеток древесины березы ( Betula papyrifera) наблюдается высокое относительное содержание сирингильных единиц, а лигнины сложной срединной пластинки и стенок сосудов богаты гваяцильными единицами. В еловых ( Picea abies) трахеидах лигнин вторичной стенки содержит в 2 раза больше фенольных ОН-групп по сравнению с лигнином сложной срединной пластинки. [6]
Из размолотой древесины лигнин извлекают водным диоксаном. Выход ЛМР составляет примерно половину всего лигнина хвойной древесины ( например, еловой) и несколько выше для древесины лиственных пород. В состав ЛМР входят часть лигнина срединной пластинки и значительная часть лигнина вторичной стенки. [7]
Исследованиями показано, что при действии сульфатного щелока на древесину сначала удаляется большая часть лигнина, находящегося в межклеточном веществе, а затем уже из первичной и вторичной стенок волокон. В этом случае остаточный лигнин распределяется как бы равномерно по всему поперечному сечению волокна и содержание лигнина во вторичной стенке будет относительно более высоким, что, по-видимому, связано с набуханием волокна под действием щелочи, делающим лигнин менее доступным для реагентов. При сульфитной варке лигнин вторичной стенки удаляется одновременно с лигнином срединной пластинки. Поэтому в отличие от волокон сульфатной целлюлозы он не распределяется в них равномерно, а концентрируется в наружных и, в меньшей степени, во внутренних слоях клеточных оболочек, так как наружные слои древесного волокна более сильно лигнифи-цированы, чем внутренние. Такое распределение лигнина в во-локнах сульфатной и сульфитной целлюлоз обусловливает и различную доступность его к действию реагентов. Так, при отбелке сульфитной целлюлозы белящий реагент легко проникает к лигнину и уже в первой стадии отбелки расходуется примерно 2 / з требуемого количества хлора. В случае сульфатных целлюлоз большая часть лигнина в стенках волокон вначале недоступна реагентам. [8]
Исходные монолигнолы включаются в протолигнин в том же порядке, в каком они образуются в ходе биосинтеза - сначала - кумаровый, затем конфериловый спирт и, наконец, синаповый. Поэтому гваяцильный лигнин хвойных неоднороден: лигнин срединной пластинки содержит большую долю Н - единиц, чем лигнин вторичной стенки, тогда как S-еди-ницы обнаруживаются в лигнине вторичной стенки ближе к полости. Состав гваяцил-сирингильного лигнина лиственных подчиняется этой же закономерности, но, соответственно, содержит меньшую долю Н - единиц ( только в лигнине срединной пластинки) и большую долю S-единиц. [9]
Терашима с сотрудниками на основании исследований, проведенных в последнем десятилетии, приходит к заключению, что протолигнин в древесине нельзя считать полностью хаотическим полимером - результатом случайной сополимеризации смеси различных монолигнолов. Лигнин образуется в присутствии и с участием полисахаридов в биологически регулируемом процессе, тесно связанном с ходом формирования ультраструктуры лигнифицированной клеточной стенки в целом. Неизбежное следствие такого протекания процессов отложения слоев клеточной стенки и их одревеснения - гетерогенность лигнина в древесине. В хвойных деревьях различаются по составу лигнины срединной пластинки и вторичной стенки, а в лиственных деревьях существуют дополнительно различия между лигнинами волокон и сосудов. Следует подчеркнуть, что образованию лигнина предшествует отложение полисахаридов - целлюлозы в виде микрофибрилл, пектиновых веществ и гемицеллюлоз разного типа для каждой стадии отложения лигнина. [10]
Страницы: 1