Генетический материал
Cтраница 1
Генетический материал обладает такими универсальными свойствами, как дискретность, непрерывность, линейность и относительная стабильность, выявляемыми в ходе генетического анализа. [1]
Генетический материал самого фага обозначен черной и белой палочками. Кроме того, в бактерию вводятся два участка бактериальных хромосом, один из которых содержит аллель X локуса, не поддающегося анализу в условиях этого опыта. При делении хромосом в районах 1, 2 или 3, отмеченных стрелками, может произойти перекрест. Очевидно, что в результате этого аллель В, который был введен в клетку при трансдукции, войдет в состав одной из двух новых бактериальных хромосом. [2]
Генетический материал был сформирован в хромосомах. [3]
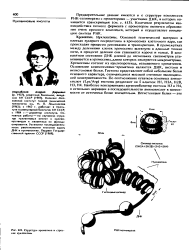 |
Структур хроматин, ние нуклеосомы. [4] |
Осиоаной генетический материал в клетках эукариот сосредоточен в хромосомах клеточного ядра, где происходят процессы репликации и транскрипции. В промежутках между делениями клеток хромосомы вытянуты в длинные тонкие нити, в процессе деления они становятся короче и толще. В компактном состоянии ДНК длиной до нескольких сантиметров упаковывается в хромосомы, длина которых измеряется микрометрами. Хромосомы состоят из нуклеопротеида, называемого хроматином. Основными компонентами хроматина являются ДНК, гистоны и иегистоновые белки. Гистоны представляют собой небольшие белки основного характера, отличающиеся высокой степенью эволюционной консервативности. Наиболее консервативны аргининбогатые гистоны НЗ и Н4, в остальных вариации а последовательности аминокислот в зависимости от источника более значительны. [5]
Генетический материал любой клетки представлен ДНК, информационные свойства которой определяются специфической последовательностью четырех нуклеотидов в полинуклеотидной цепи. Полуконсервативный механизм репликации ДНК, в результате которого из одной родительской двухцепочной молекулы образуются две дочерние молекулы, содержащие по одной родительской и одной вновь синтезированной комплементарной полинуклеотидной цепи, наилучшим образом обеспечивает идентичность исходной и синтезированных молекул и, следовательно, сохранность видоспецифической наследственной информации в ряду поколений клеток и организмов ( см. гл. [6]
 |
Ретиналь ( А и предполагаемая организация. [7] |
Генетический материал экстремальных галофилов представлен в виде основной и сателлитных ДНК. Последние составляют от 11 до 30 % всей содержащейся в клетках ДНК и состоят из замкнутых кольцевых молекул. Высокий уровень сателлитных ДНК - уникальная черта организации генетического материала экстремальных галофилов, значение которой пока не ясно. Предполагается, что сателлитные ДНК - не эписо-мы, а составная часть генома этих бактерий. [8]
Их генетический материал расположен в линейной последовательности, и на основе рекомбинаций признаков, которые влияют на выражение симптомов, можно построить генетические карты ( см. гл. [9]
Способность генетического материала, ДНК, к самовоспроизведе-нию ( репликации) лежит в основе размножения живых организмов, передачи наследственных свойств из поколения в поколение и развития многоклеточного организма из зиготы. [10]
Способность генетического материала, ДНК, к самовоспроизведению ( репликации) лежит в основе размножения живых организмов, передачи наследственных свойств из поколения в поколение и развития многоклеточного организма из зиготы. [11]
 |
Структурная схема управления генетического поиска. [12] |
Перераспределение генетического материала на основе генетических операторов и их модификаций выполняется внутри каждого строительного блока. За счет этого уменьшается размер хромосом в популяции и сокращается время реализации основного алгоритма. Например, в хромосоме рь 2, 3, 4, 5, которая является строительным блоком для графа G ( рис. 6.71, а), определим точку мутации между 3 и 4 геном. [13]
Перетекание генетического материала в плазмидные геномы и обратно строго доказано. Модулярная природа плазмидной эволюции может служить объяснением разнообразия катаболи-ческих фенотипов, связанных с этой группой плазмид. [14]
Перенос генетического материала строго ориентирован: разрыв копии хромосомы и передача ДНК происходит в локусе 0 в пределах полового фактора. Скорость переноса в одинаковых условиях для определенного штамма является постоянной. Обычно всей хромосоме не удается перейти в клетку-реципиент, так как контакт клеток очень нестабилен и часто прерывается до завершения перехода. [15]
Страницы: 1 2 3 4