Пурпурные зеленые бактерия
Cтраница 2
Всего сейчас описано около пятидесяти видов пурпурных и зеленых бактерий. Многи & чиз них выделены в виде чистых культур, или монокультур. В морфологическом отношении фото-трофные бактерии проявляют значительное разнообразие. Некоторые фототрофные бактерии, особенно часто в природных условиях, образуют различные, иногда весьма характерные скопления ( агрегаты) клеток в виде цепочек и пластинок, нередко заключенных в слизь. Размеры фототрофных бактерий также весьма различные, от 1 - 2 мкм в диаметре до 50 мкм и более в длину. Наиболее крупные формы встречаются среди пурпурных серобактерий. К ним относятся такие организмы, как Chromatium okenii и Thiospirillum jenense. Есть подвижные и неподвижные виды. У боль-пганства движение обусловлено наличием одного или нескольких жгутиков, расположенных полярно. Лишь у одного вида пурпурных бактерий ( Rhodomicrobium vannielii) жгутикование перитрихиальное. В зависимости от направления движения эти реакции могут быть положительными или отрицательными. Большинство фототрофных бактерий размножается бинарным делением. [16]
В процессе восстановления С02 на свету пурпурными и зелеными бактериями не происходит выделения кислорода. [17]
Всеми вышеуказанными работами было показано, что пурпурные и зеленые бактерии способны осуществ - лять восстановление углекислоты на свету, но в отличие от фотосинтеза зеленых растений они при этом кислорода не выделяют. [18]
Может быть, это открытие указывает на то, что пурпурные и зеленые бактерии - предшественники зеленых растений, реликты того времени, когда земная кора была менее устойчива в химическом отношении, чем сейчас, и сероводород, сера, а может быть, и свободный водород имелись в гораздо большем количестве. [19]
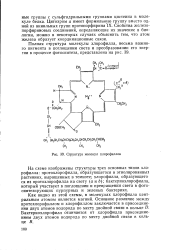 |
Структура молекул хлорофиллов. [20] |
На схеме изображены структуры трех основных типов хлорофилла: протохлорофилла, образующегося в этиолированных растениях, выращенных в темноте; хлорофилла, образующего ся из протохлорофилла на свету ( аи Ь); бактериохлорофилла, который участвует в поглощении и превращении света в фото-синтезирующих пурпурных и зеленых бактериях. [21]
Ионы аммония подавляют синтез нитрогеназы. У пурпурных и зеленых бактерий под влиянием этих ионов уменьшается также активность уже синтезированного фермента. В регуляции образования нитрогеназы большую роль, очевидно, играет глу-таминсинтетаза. Глутаминсинтетаза и глутаматсинтаза нужны бакте-риям для включения ионов аммония в органические соединения в том случае, если эти ионы присутствуют лишь в низкой концентрации. Эта система обладает высоким сродством к ионам аммония и поддерживает их концентрацию в клетке на низком уровне. Повышение концентрации ионов аммония в окружении клетки ( а тем самым и внутри клетки) подавляет образование глутаминсинтетазы, а в результате-и нитрогеназы. [22]
 |
Пути окисления соединений серы фототроф-нымн бактериями АФС - адепилилсульфат. [23] |
Для других видов, даже выделенных из очень соленых водоемов, она более низкая. И для пурпурных и зеленых бактерий, выделенных из соленых водоемов, присутствие NaCl обязательно. [24]
В настоящее время фототрофные бактерии широко используют для исследования фотосинтеза в различных аспектах, особенно начальных стадий, поскольку они удобны для изучения этого сложного вопроса. Кроме того, пурпурные и зеленые бактерии интересны для выяснения организации фотосинтезирующего аппарата, путей биосинтеза пигментов, метаболизма углерода, эволюции фотосинтеза и фотосинте-зирующих форм. Привлекают они к себе внимание и в связи с другими биологическими проблемами, в частности фиксацией молекулярного азота, а также круговоротом углерода и серы в природе. [25]
Чувствительность микробов к свету различна. Для водорослей, пурпурных и зеленых бактерий, синтезирующих органическое вещество посредством ассимиляции углекислоты, свет является основным условием существования. Известно, что в процессе фотосинтеза флора поверхностных слоев воды использует углерод из СО2 и выделяет в окружающую среду кислород. Поэтому свет оказывает большое влияние на окислительные процессы. [26]
Интересным макрообразованием является циано-бактериалъный мат. В его состав входят цианобактерии, пурпурные и зеленые бактерии, а также сульфатредукторы. [27]
Три основных фактора определяют распространение фототроф-ных эубактерии в природе: свет, молекулярный кислород и питательные вещества. Потребности в разных частях солнечного спектра для фотосинтеза определяются набором светособирающих пигментов. Пурпурные и зеленые бактерии часто развиваются в водоемах под более или менее плотным поверхностным слоем, состоящим из цианобактерий и водорослей, эффективно поглощающих свет до 750 нм. [28]
В условиях интенсивного роста фототрофных бактерий большая часть первичных продуктов ассимиляции углекислоты и органических соединений используется на синтез белков и других важнейших компонентов клеток. Но при задержке роста культур в результате недостатка фосфора и особенно азота происходит образование в больших количествах запасных веществ. И пурпурные и зеленые бактерии могут накапливать в качестве запасных продуктов полисахариды типа гликогена. Накопление того или другого запасного соединения зависит не только от особенностей организма, но и от источника углерода. Полисахариды преимущественно накапливаются пурпурными бактериями при культивировании их на минеральных средах, а также в присутствии пирувата, пропионата, малата или сукцината. При использовании бутирата или ацетата, особенно в отсутствие бикарбоната, происходит накопление почти исключительно поли-р-оксибутирата, так как синтез его начинается с конденсации двух молекул ацетил - КоА с образованием аце-тоацетил - КоА и далее р-оксибутирата. [29]
То, как эта возможность реа-лизовывалась, доказывает наличие нескольких типов фотосинтеза, осуществляемого разными группами эубактерий, энергетический метаболизм которых полностью или частично основан на использовании энергии света. Фотосинтезирующие эубактерий представлены пурпурными и зелеными бактериями, гелио-бактериями, цианобактериями1 и прохлорофитами. [30]
Страницы: 1 2 3