Зеленые нитчатые бактерия
Cтраница 1
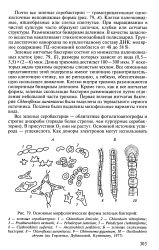 |
Основные морфологические формы зеленых бактерий. [1] |
Зеленые нитчатые бактерии состоят из множества палочковидных клеток ( рис. 79, Б), размеры которых зависят от вида ( 0 5 - 5 5) х ( 2 - 6) мкм. Длина трихомов достигает 100 - 300 мкм. У некоторых видов трихомы окружены слизистым чехлом. Все описанные представители этой подгруппы имеют типичную грамотрицатель-ную клеточную стенку, но не ригидную, а гибкую, обеспечивающую скользящее движение. Клетки внутри трихома размножаются поперечным бинарным делением. Кроме того, как и все нитчатые формы, зеленые скользящие бактерии размножаются путем отделения небольшой части трихома. Первая зеленая нитчатая бактерия Chloroflexus aurantiacus была выделена из термального серного источника. Позднее были выделены мезофильные варианты этого вида. [2]
У пурпурных и зеленых нитчатых бактерий, у которых функционирует только светозависимый циклический электронный транспорт, нет надобности в заполнении электронной вакансии в молекуле хлорофилла. В то же время проблема получения фотохимическим путем восстановителя не решена. [3]
Физиолого-биохимическая характеристика зеленых нитчатых бактерий основана главным образом на данных, полученных для разных штаммов Chloroflexus aurantiacus, обнаруживших значительное метаболическое разнообразие. На свету он растет в аэробных и анаэробных условиях в присутствии разнообразных органических соединений: Сахаров, спиртов, органических кислот и аминокислот. Некоторые штаммы этого вида способны к анаэробному фотоавтотрофному росту, используя Н2 или H2S в качестве донора электронов. Окисление H2S приводит к образованию молекулярной серы и отложению ее в среде в виде аморфной массы. Молекулярная сера в очень незначительной степени затем окисляется до сульфата. Хемогетеротрофный рост также возможен в аэробных и для отдельных штаммов в анаэробных условиях. [4]
Первый представитель зеленых нитчатых бактерий Chloroflexus aurantiacus был выделен из термального источника, где рос, формируя пленку толщиной несколько миллиметров. Позднее термофильные штаммы этого вида были найдены во многих нейтральных и щелочных горячих источниках с температурой от 45 до 75 С, где условия, как правило, микроаэробные. Часто Chloroflexus образует смешанные популяции с термофильными цианобакте-риями рода Synechococcus. Вскоре из природных слоев пресных озер были выделены мезофильные аналоги Chloroflexus с оптимальной температурой роста 20 - 25 С. [5]
Отметим также, что у зеленых нитчатых бактерий Chloroflexus было показано, что ассимиляция углекислоты осуществляется не только в цикле Кальвина, но и необычными путями - в 3-гидрокси-пропионатном цикле и / или восстановительном цикле дикарбо-новых кислот. [6]
Циклическим электронным транспортом у фотосинтезирующих эубактерий не исчерпываются все возможные пути переноса электронов. Электрон, оторванный от первичного донора реакционного центра, может по цепи, состоящей из других переносчиков, не возвращаться к молекуле хлорофилла, а передаваться на такие клеточные метаболиты, как НАД ( Ф) или окисленный ферредок-син, которые используются в реакциях, требующих восстановителя. Таким образом, электрон, покинувший молекулу хлорофилла, выводится из системы. Возникает однонаправленный незамкнутый электронный поток, получивший название нециклического пути переноса электронов. У пурпурных и зеленых нитчатых бактерий функционирует только циклический светоза-висимый поток электронов. У остальных групп эубактерий фото-индуцируется как циклический, так и нециклический перенос электронов, при этом у зеленых серобактерий и гелиобактерий оба пути электронного транспорта связаны с функционированием одной фотосистемы, а у цианобактерий и прохлорофит циклический перенос электронов зависит от активности фотосистемы I, a для нециклического потока электронов необходимо функционирование обеих фотосистем. Поток электронов по цепи переносчиков на определенных этапах сопряжен с направленным перемещением протонов через мембрану, что приводит к созданию протонного градиента, используемого для синтеза АТФ. [7]
Страницы: 1