Плазматическая мембрана
Cтраница 1
 |
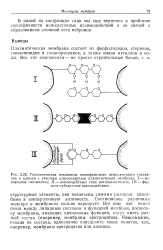
Модель структуры плазматической мембраны. [1] |
Плазматическая мембрана состоит из двойного липидного слоя. Гидрофобные концы молекул фосфолипидов и триглицеридов направлены внутрь, а гидрофильные головки - наружу. Благодаря гидрофобным взаимодействиям между остатками жирных кислот, входящих в состав липидов, и электростатическому взаимодействию между гидрофильными головками мембрана стабилизируется. В двойной слой липидов встроены белки - так называемые интегральные белки мембран. Они плавают в этом слое, будучи погружены в него частично, или же пронизывают его насквозь. Некоторые мембраны, по-видимому, с одной или с обеих сторон покрыты сетью вытянутых белковых молекул. [2]
Плазматическая мембрана играет важнейшую роль в обмене веществ. Она служит осмотическим барьером клетки и контролирует как поступление веществ внутрь клетки, так и выход их наружу. В мембране имеются механизмы активного транспорта и системы субстрат-специфичных пермеаз. По-видимому, липидная пленка элементарной мембраны пронизана мостиками ( или каналами) из белков, и именно эти белки служат порами, через которые осуществляется регулируемый транспорт веществ. [3]

Плазматическая мембрана состоит из двойного липидного слоя. Гидрофобные концы молекул фосфолипидов и триглицеридов направлены внутрь, а гидрофильные головки - наружу. [5]
 |
Состав мембран Micrococcus luteus ( lysodeikticus и фототрофных бактерий.| Модель структуры плазматической мембраны. [6] |
Плазматическая мембрана играет важнейшую роль в обмене веществ. Она служит осмотическим барьером клетки и контролирует как поступление веществ внутрь клетки, так и выход их наружу. В мембране имеются механизмы активного транспорта и системы субстрат-специфичных пермеаз. По-видимому, липидная пленка элементарной мембраны пронизана мостиками ( или каналами) из белков, и именно эти белки служат порами, через которые осуществляется регулируемый транспорт веществ. [7]
Плазматическая мембрана вместо глицериновых эфиров жирных кислот содержит глицериновые эфиры с С20 ( фитанил) - и С4о ( бифи-танил) - изопреноидными алкилами. [8]
Плазматическая мембрана состоит из фосфолипидов, стеринов, гликолипидов и гликопротеинов, а также ионов металлов и воды. [10]
Плазматические мембраны лимфоцитов ( содержащие 20 мг общего белка) экстрагировали 5 мл 1 % - ного дезоксихолата, после чего растворимую фракцию ( содержащую 85 % мембранного белка) фильтровали через колонку с аффинным сорбентом. При элюировании 1 % - ным раствором дезоксихолата до нулевой базовой линии ( при 280 нм) отмывали примерно 75 % сорбированного белка. Рецепторные гликопротеины ( составляющие 5 % от фракции мембранных белков) элюировали раствором метил-сс-с-глюкопиранозида ( 20 мг / мл) в 1 % - ном дезоксихолате. Элюат подкисляли 2 % - ной уксусной кислотой ( добавляя 0 1 объема элюата), выпавший осадок промывали тремя порциями по 10 мл 95 % - ного этанола для удаления моносахарида и дезоксихолата, а затем высушивали в токе воздуха. [11]
Плазматические мембраны нейронов и мембраны некоторых не нейрональных клеток содержат специфические рецепторы ( рецепторы NGF), которые связывают NGF вначале с низким, а затем с высоким сродством. Было показано, что рецепторы с высоким сродством образуют кластеры и вместе со связанным NGF попадают в клетку при эндоцитозе и транспортируются внутри клетки частично к лизосомам ( где происходит их деградация), частично к ядру. При их поглощении нервным окончанием рецептор и NGF переносятся путем ретроградного аксонального транспорта. Подобные процессы могут происходить и при других типах гормональной регуляции и поэтому NGF служит своеобразной моделью гормонов и факторов роста. Механизм действия NGF в клетке не изучен. [12]

Плазматические мембраны других клеток имеют более сложное строение. На внешней поверхности клеток во многих плотных тканях присутствует еще один важный гликопротеин - мбронек-тин ( разд. [14]
Плазматические мембраны соседних клеток переходят непосредственно одна в другую, выстилая поры. Через просвет каждой поры переходит из клетки в клетку и агранулярный эндоплазма-тический ретикулум. Такая система упрощает связи и координацию между отдельными растительными клетками, поскольку ионам и молекулам не приходится преодолевать на своем пути плазматическую мембрану. Вирусы способны использовать поры в клеточных стенках и могут переходить из клетки в клетку по плазмодесмам. [15]
Страницы: 1 2 3 4 5