Мессенджер
Cтраница 1
Мессенджер и Скотт [93] предварительно подвергали кристаллизации невулканизованный каучук, а затем изучали его поведение при растяжении. [1]
Для этого высвобождения химический вторичный мессенджер, вероятно Са2 и / или cGMP, должен пройти путь от мембраны дисков, содержащей родопсин, до плазматической мембраны клетки палочки. Свет стимулирует фосфорилирование родопсина - процесс, который может быть связан с адаптацией сетчатки к изменяющимся световым условиям. [2]
К внутриклеточной системе мессенджеров относят также производные фосфолипидов мембран эукариотических клеток, в частности фосфорили-рованные производные фосфатидилинозитола. Эти производные освобождаются в ответ на гормональный сигнал ( например, от вазопрессина или тиротропина) под действием специфической мембраносвязанной фосфо-липазы С. В результате последовательных реакций образуются два потенциальных вторичных мессенджера-диацилглицерол и инозитол-1 4 5-три-фосфат. [3]
Биологические эффекты этих вторичных мессенджеров реализуются по-разному. Действие диацилглицерола, как и свободных ионов Са2, опосредовано через мембраносвязанный Са-зависимый фермент протеин-киназу С, которая катализирует фосфорилирование внутриклеточных ферментов, изменяя их активность. Инозитол-1 4 5-трифосфат связывается со специфическим рецептором на эндоплазматическом ретикулуме, способствуя выходу из него ионов Са2 в цитозоль. [4]
В большинстве случаев Са2 как своеобразный мессенджер влияет на клеточные процессы не непосредственно, а в основном через активацию им внутриклеточного рецептора - белка кальмо-дулина. Этот белок был впервые обнаружен в тканях мозга американским исследователем В. Позднее кальмодулин был найден практически во всех растительных и животных тканях. Кальмодулин - очень кислый белок ( рН 3 9 - 4 3) благодаря тому, что около 30 % входящих в его состав остатков приходится на долю аспарагиновой и глутаминовой кислот. [5]
Таким образом, представленные данные о вторичных мессенджерах свидетельствуют о том, что каждой из этих систем посредников гормонального эффекта соответствует определенный класс протеинкиназ, хотя нельзя исключить возможности существования тесной связи между этими системами. Активность протеинкиназ типа А регулируется цАМФ, протеин-киназы G - цГМФ; Са2 - кальмодулинзависимые протеинкиназы находятся под контролем внутриклеточной [ Са2 ], а протеинкиназа типа С регулируется диацилглицеролом в синергизме со свободным Са2 и кислыми фосфолипидами. Повышение уровня какого-либо вторичного мес-сенджера приводит к активации соответствующего класса протеинкиназ и последующему фосфорилированию их белковых субстратов. В результате меняется не только активность, но и регуляторные и каталитические свойства многих ферментных систем клетки: ионных каналов, внутриклеточных структурных элементов и генетического аппарата. [6]
Отдельно запротоколировано votum separatum доктора физики Арчибальда Мессенджера, который заявил, что рассказанное Бертоном могло, по его мнению, происходить в действительности и нуждается в добросовестном изучении. [7]
Молекулярные механизмы передачи гормонального сигнала и роль вторичных мессенджеров ( посредников) в реализации гормонального эффекта подробно изложены в конце данной главы. [8]
 |
Аденилатциклазный путь передачи гормонального сигнала. [9] |
Общим фундаментальным механизмом, посредством которого реализуются биологические эффекты вторичных мессенджеров внутри клетки, является процесс фосфорилирования - дефосфорилирования белков при участии широкого разнообразия протеинкиназ, катализирующих транспорт концевой группы от АТФ на ОН-группы серина и треонина, а в ряде случаев-тирозина белков-мишеней. Процесс фосфорилирования представляет собой важнейшую посттрансляционную химическую модификацию белковых молекул, коренным образом изменяющую как их структуру, так и функции. В частности, он вызывает изменение структурных свойств ( ассоциацию или диссоциацию составляющих субъединиц), активирование или ингибирование их каталитических свойств, в конечном итоге определяя скорость химических реакций и в целом функциональную активность клеток. [10]
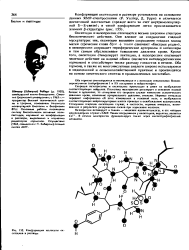 |
Конформации молекулы окситоцина в растворе. [11] |
Вазопресснн активирует в тканях - мишенях аденилатциклазу, и его вторым мессенджером служит с AMP. В печени вазопресснн функционирует также через ннозиттрифосфатный механизм. [12]
Наряду с экспериментами, типичными для планетных исследований, в задачах миссий Мессенджер и БепиКоломбо исследователи выделяют изучение таких особенностей планеты, которые присущи только Меркурию. Цель предлагаемой работы - перечислить несколько подобных фундаментальных проблем и более детально остановиться на одной из них. Так или иначе, они входят в задачи обеих миссий. [13]
Этот сигнал рецепторы трансформируют в изменение концентраций внутриклеточных посредников, получивших название вторичных мессенджеров, уровень которых определяется активностью ферментов, катализирующих их биосинтез и распад. [14]
Физиологическая регуляция происходит не только путем синтеза и циркуляции активных соединений и химических мессенджеров. Она должна осуществляться также возможностью инактивации циркулирующих сигналов. На уровне фосфорилированных белков фосфопро-теинфосфатазы гарантируют обратимость белкового фосфорили-рования. [15]
Страницы: 1 2 3