Механизм - инверсия
Cтраница 1
Механизм инверсии [883] понятен из приведенной схемы, он включает на промежуточных стадиях комплексы аллильного типа. [1]
Механизм инверсии смачивания связан с определенной ориентацией молекул ПАВ в адсорбционном слое. Если твердая поверхность первоначально гидрофильна, то адсорбированные молекулы взаимодействуют своими полярными группами с поверхностью, а неполярными цепями обращаются наружу, вследствие чего твердая поверхность становится гидрофобной. Например, при погружении стеклянной пластинки в раствор стеариновой кислоты в октане или бензоле на поверхности пластинки образуется монослой стеариновой кислоты. Адсорбированные молекулы кислоты на пластинке ориентируются неполярными цепями наружу, придавая поверхности гидрофобные свойства. [2]
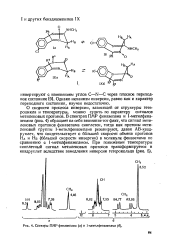 |
Спектры ПМР феназепама ( а и 1-метилфеназепама ( б. [3] |
Однако механизм инверсии, равно как и характер переходного состояния, изучен недостаточно. [4]
Каждый механизм инверсии спина дает только ему присущий стереохи-мический результат. Следовательно, стереоспецифичность химических реакций является результатом не только электронных особенностей синглетных реакций, но и результатом спин-орбитальных взаимодействий в процессе реакций триплетных состояний. [5]
В целом механизм инверсии, предложенной Пар-неллом и Богеменом, представляется более обоснованным, хотя искривление изотермы теплопроводность - состав, по-видимому, также имеет важное значение. [6]
С точки зрения единообразия механизмов инверсии и методов накачки лазеры классифицируют по типу фазового состояния активного вещества. [7]
Рацемизация оптически активных изомеров может происходить по механизму инверсии. Поэтому возможно и разделение его оптических изомеров и их рацемизация. Однако барьер инверсии для большинства аминов низок ( ж40 кДж / моль для пропилфенилэтиламина и ж25 кДж / моль для аммиака), и оптические изомеры не могут быть разделены, потому что рацемизация осуществляется быстрее, чем разделение. Для идентификации оптических изомеров в рацематной смеси и изучения скорости взаимопревращения энантиомеров следует обратиться к спектроскопическим методам. Если энергетический барьер инверсии лежит в интервале 20 - 100 кДж / моль, лучшим методом изучения изомеров является ядерный магнитный резонанс. Переходное состояние, возникающее в ходе атомной инверсии, будет отвечать плоскому окружению центрального атома, а энергетический барьер инверсии будет зависеть от того, насколько легко молекулу можно вывести из ее основного состояния. Меньшие углы между связями в фосфинах, чем в соответствующих аминах, требуют больше энергии для перехода в плоское состояние, следовательно, для фосфинов характерна большая оптическая устойчивость. Наличие электроноак-цепторных заместителей повышает высоту барьера, а электроно-донорные группы уменьшают его. [8]
Обнаружение оптически активного замещенного ( агепе) Сг ( СО) 3 [102] позволяет проверить этот механизм инверсии для обмена арена. [9]
Сравнение констант & кл и & тун, рассчитанных по уравнениям (1.37) и (1.44), показывает, что туннелирование играет существенную роль в механизмах инверсии молекулы аммиака и внутреннего вращения в этане. Но уже в 1 2-дихлорэтане, благодаря большей массе молекулы и большей высоте барьера, & кл при комнатных температурах значительно превышает / тун. Роль туннелирования, естественно, возрастает при понижении температуры. [10]
В табл. 111 - 10 приведены данные времени жизни капель масла в воде ( т) и воды в масле ( тв) в системе без соли и в системе, насыщенной NaCl, для всех исследованных ПАВ. Для выявления механизма инверсии проделаны следующие опыты. В системе без соли тм тв, при этом образуется прямая эмульсия. [11]
И снова структура С4в не может быть активированным комплексом для любой реакции замещения, поскольку он может распадаться по четырем эквивалентным направлениям. Эта структура лежит на половине пути между двумя структурами D 3h меньшей энергии, которые представляют собой активированные комплексы для механизма инверсии. Она лежит также в центре между четырьмя структурами симметрии Cs меньшей энергии, которые превращаются одна в другую при более низкой энергии, чем структура С4 в, и дают сохранение конфигурации. [12]
Некоторые D-аминокислоты при включении их в рацион взамен соответствующих L-изомеров могут поддерживать рост животных и размножение микроорганизмов. В большинстве случаев эти D-изомеры, очевидно, превращаются в L-аминокислоты путем окислительного дезаминирования с образованием а-кето-кислот и последующего реаминирования в результате переами-нирования. Следует подчеркнуть, что такой механизм инверсии D-аминокислот в организме нельзя считать полностью доказанным; однако эти представления находятся в согласии с данными, свидетельствующими о возможности обеспечения роста животных и микроорганизмов при помощи а-кетоаналогов некоторых аминокислот, а также с существованием и широким распространением оксидазы D-аминокислот и различных транс-аминаз. Наблюдения над ростом молодых крыс показали, что вместо D-аминокислот, которые заменяют соответствующие L-аминокислоты, можно вводить в организм и соответствующие а-кетопроизводные. Вместе с тем в некоторых случаях а-кето-кислоты ( например, а-кетоаналоги лейцина и изолейцина) обеспечивают рост молодых животных, тогда как соответствующие им D-аминокислоты не используются растущим организмом; эти данные позволяют предполагать, что скорость окислительного дезаминирования D-аминокислот не всегда достаточно высока, чтобы обеспечить рост животных. [13]
Страницы: 1