Механизм - активность
Cтраница 1
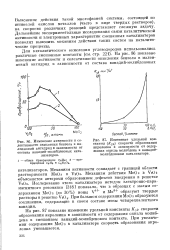
Механизм активности совпадает с границей области растворимости МоОз в VzOs. Механизм действия МоОз в VaOs объясняется авторами образованием дефектов внедрения в решетке УЮб. При большом содержании МоО з образуются соединения, содержащие в своем составе ионы четырехвалентного ванадия. [2]
Картина механизма ксантиноксидазной активности, которая вырисовывается главным образом по результатам опытов, проведенных методом ЗПР, состоит в том, что фермент содержит два независимых каталитических центра. Электрон переносится на конечный электронодонорный флавино-вый центр непосредственно или при участии железосерусодержаще-го фрагмента. [3]
В механизме бласттрансформирующей активности, по-видимому, принимают участие лимфокины. [4]
Показано, что механизм гипотензивной активности каптоприла связан с его способностью снижать уровень ангиотензина II и аккумулировать содержание в организме брадикинина, обладающего сосудорасширяющим действием. [5]
Особенно важную роль в системе механизмов информационной активности играют познавательные интересы студентов. Определяя структуру информационных интересов, они выполняют роль своеобразного поведенческого ориентира. Причем познавательный интерес выступает как внутренний источник, как побудительная сила информационной активности студента. Маркс-за что человек борется, связано с его интересом... Поэтому интересы нужно рассматривать как главный внутренний стимулятор активности. И чем разностороннее и устойчивее познавательные интересы студентов, тем многообразнее по форме и богаче по содержанию будет их познавательная активность, тем успешнее будет совершаться процесс подготовки квалифицированных специалистов. [6]
При управлении эффективностью производства энергии для сокращения мышц и других метаболических и органических процессов, связанных с производством энергии, наиболее существенную приспособительную роль играют механизмы мо-орной активности и моторной интеграции. Актин и миозин - это палочкообразные молекулы ( нити), которые располагаются параллельно в мышечной клетке и связаны между собой скрещивающимися мостиками, которые образуются в присутствии кальция путем соединения головной части миози-новой нити с определенным местом на соседней нити актина. В процессе сокращения меняется структура мостиков и запускается повторяющийся сократительный цикл, вызывающий попеременное растяжение нитей актина и миозина и в результате укорачивание мышцы. [7]
Теория пересыщения не отражает многих сторон активности катализатора; кроме общих указаний на три достоверных источника пересыщений ( которые обстоятельно изучены и подтверждены опытом), она не вскрывает щикаких механизмов активности. Обобщив большой экспериментальный материал определенной области явлений, она указывает практические пути создания высоко пересыщенных систем, являющихся, как правило ( но е всегда), активными катализаторами - в этом е достоинство. В связи с предшествующим опытом, обобщенным и рационально направленным на решение новых практических задач - ее материальная основа. [8]
Исследования в области физиологии опорно-двигательного аппарата сосредоточены на изучении нейронных механизмов движения, разработке методов познания биомеханики движений человека с использованием ЭВМ, моделировании движений многозвенных кинематических цепей. Ведутся также работы, посвященные изучению механизмов позной активности и выяснению механизмов сенсорного обеспечения движений. [9]
Механизмы действия этих соединений связывают с непрямым симпатоми-метическим эффектом. Недавнее исследование гидролиза сиднокарба и сиднофена показало, что в окислительных условиях происходит образование оксида азота и потому, вероятно, высвобождение оксида азота в организме при применении этих препаратов. Установление вклада высвобождающегося NO в механизм психотропной активности сиднокарба и сиднофена требует специальных биохимических и фармакологических исследований. [10]
В том случае, когда нейрон под влиянием двух разных пейсмекерных потенциалов генерирует два независимых ПД, возникает возможность проверить предположение о том, что адаптация может быть связана с локальными процессами изменения сопротивления или подключения источника тока в каждом пейсмекерном локусе. В таком нейроне подача деполяризующего тока вызывает сильную реакцию в виде учащения одного ПД, в то время как действие тока на другой ПД минимально. Более внимательный анализ показывает, что в первый момент ток действует на оба ПД, но затем в одном локусе быстро развивается адаптация, и ток перестает влиять на частоту ПД, в то время как в другом локусе адаптация минимальна и на всем протяжении действия тока частота ПД сохраняется увеличенной. При выключении деполяризующего тока в том локусе, где адаптация отсутствовала, частота сразу же возвращается к исходному значению. В другом локусе, где адаптация к деполяризующему току была выражена сильнее, выключение деполяризующего тока приводит к тому, что частота ПД снижается, указывая на эффект работы активного транспорта ионов, эквивалентной инъекции слабого гиперполяризующего тока. Гиперполяризующий ток, подаваемый через микроэлектрод, подавляет механизм пейсме-керной активности на обоих пейсмекерных участках. Затем, в связи с преимущественным развитием адаптации к току в одном локусе, частота одного ПД приближается к исходному значению. В другом локусе адаптация не наступает и частота этого ПД остается сниженной на всем протяжении действия гиперполяризации. В соответствии с представлением об активном транспорте как одном из источников адаптации при выключении гиперполяризующего тока в том локусе, где развилась адаптация, наступает учащение ПД. В том локусе, где адаптация не наступала, частота ПД после выключения гиперполяризации сразу же возвращается к норме. [11]
Страницы: 1