Зародышевые мешка
Cтраница 1
Зародышевые мешки также чувствительны к подобным изменениям, но в меньшей степени, чем пыльцевые зерна. Таким образом, разумно допустить, что частичная стерильность, нередко наблюдаемая у перекрестноопыляющихся видов, вызвана изменчивостью структуры хромосом. Исследования хромосом ржи, проведенные А. Праккеном, показали, что подобная изменчивость действительно встречается у этого вида. Из 167 растений, взятых из популяций, принадлежащих к двум различным сортам ржи, 6 % были гетерозиготны по транслокациям и инверсиям или имели другие значительные изменения хромосом. Среди остальных растений у многих наблюдались менее резкие нарушения мейоза, причины которых не удалось проанализировать достаточно глубоко. [1]
Зародышевые мешки обычно лучше выносят отклонения от нормального числа хромосом, но, как правило, плодовитость по женской линии также снижается. [2]
Это касается и женских половых клеток: зародышевые мешки с несбалансированным хромосомным набором не способны функционировать. [3]
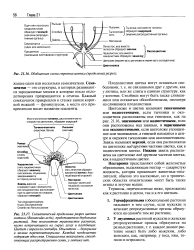
Семязачатки - это структуры, в которых развиваются зародышевые мешки и которые после оплодотворения превращаются в семена. Каждый семязачаток прикреплен к стенке завязи короткой ножкой - фуникулусом, а место его прикрепления носит название плаценты. [5]
У этого вида В-хромосомы ведут себя необычно: они элиминируются из большей части вегетативных тканей, сохраняясь лишь в тех частях растения, которые образуют пыльцевые зерна и зародышевые мешки. [6]
Однако чаще некоторая часть материнских клеток пыльцы ( и материнских клеток зародышевого мешка) содержит прямые транслокационные кольца, в которых лежащие рядом хромосомы направляются к одному и тому же полюсу. В результате образуются абортивные пыльцевые зерна и зародышевые мешки. Это показывает, что частота зигзагообразных и прямых колец в первой метафазе примерно одинакова. [7]
У родов растений с высокими основными числами эффект, оказываемый анеуплоидностью, также слабее, чем у родов с низким основным числом. Так, у триплоидной осины ( Рори-lus), имеющей 3 - 19 57 хромосом, анеуплоидные пыльцевые зерна и зародышевые мешки функционируют нормально; анеуплоидные дочерние растения в среднем развиваются также лучше, чем соответствующие анеуплоиды у видов с низким основным числом хромосом. Это хорошо согласуется с предположением, что высокое основное число хромосом у Populus следует рассматривать как пример вторичной полиплоидии. Иными словами, основное число хромосом у осины представляет замаскированную полиплоидию и диплоидная осина на самом деле - полиплоид, происшедший от неизвестных нам и, возможно, вымерших предков с более низкими основными числами хромосом. [8]
У настоящих гаплоидов, имеющих лишь по одной хромосоме каждого типа, мейоз протекает, конечно, весьма неправильно. Пары хромосом, состоящие из двух гомологов, соединенных хиазмами, образоваться не могут, поэтому хромосомы распределяются случайно ( фиг. Таким образом, в большинстве случаев пыльцевые зерна и зародышевые мешки получают неполный, несбалансированный набор хромосом, дающий летальный эффект. [9]
В других случаях тормозится второе деление мейоза, но при этом также в результате образуются нередуцированные пыльцевые зерна и яйцеклетки. В редких случаях возникают как пыльцевые зерна, так и зародышевые мешки, у которых число хромосом вдвое больше соматического ( фиг. [10]
У бессеменных растений каждая мегаспора прорастает в жен. Иногда клеточная перегородка закладывается только после первого деления мейоза, а после второго не образуется. Такая двуядерная клетка соответствует двум необособившимся мегаспорам и из нее развивается двухспоровый ( биспори-ческий) зародышевый мешок ( напр. Двух - и четырехспоровые зародышевые мешки считаются производными, возникшими позднее в ходе эволюции. У цветковых растений мегаспорофиллу функционально и морфологически соответствует плодолистик. [11]
Страницы: 1