Модель - популяция
Cтраница 1
Модель популяции, в которой могут возникать эпидемии, можно построить следующим образом. [1]
Модели популяции, в которых учитывается неоднородность возраста и миграция, описываются следующим образом. [2]
Эта модель популяции более устойчива по сравнению с моделями, рассмотренными в предыдущих разделах. Устойчивость выражается в том, что колебания являются затухающими, но более подробные сведения, касающиеся модели, нас интересовать не будут. Важно то, что численность популяции жертвы, с запаздыванием зависящая от плотности, регулируется хищником. Однако, если мы нанесем на график значения k, смертности, вызванной хищником за время генерации, в зависимости от логарифма плотности жертв ( рис. 10.3, Б), то четкой связи не получим, Несмотря на то, что воспользовались общепринятым способом представления зависимости от плотности ( гл. Но если те же самые точки мы соединим последовательно от поколения к поколению ( рис. 10.3, В), то увидим, что они ложатся на спираль, закрученную против часовой стрелки. Эта спиралевидная связь и характеризует зависимость от плотности с запаздыванием; из-за того, что колебания затухающие, спираль закручейа внутрь. [3]
Кинетический подход обусловливает и, можно сказать, диктует выбор определенного типа модели популяции. Как уже упоминалось, в этом случае обращаются к распределительной модели эндогенно расширяющейся биофазы. Уровень структуры, участвующий в элементарном акте и рассматриваемый неделимо, определяется позицией исследователя и смыслом конкретно решаемой задачи моделирования. [4]
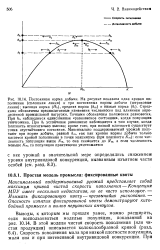
Выводы, к которым мы пришли ранее, можно расширить, если воспользоваться моделью популяции, показанной на рис. 10.14. В этой модели чистая скорость пополнения ( или чистая продукция) описывается колоколообразной кривой ( разд. Скорость пополнения мала при низкой плотности популяции, мала она и при интенсивной внутривидовой конкуренции. [6]
 |
Возможные результаты бимодального эксперимента ( ДА 1, НЕТ 0 и их вероятности ( при п - 3. [7] |
Термин случайная величина относится к такой переменной, для каждого значения которой существует определенная вероятность принятия этого значения. Теоретической схемой распределения значений случайной величины является модель популяции. Исследуемая группа соответствует распределению частоты принятия значений. Это полезный метод графического изображения данных; он заключается в составлении гистограммы распределения со значениями интересующей переменной вдоль горизонтальной оси и частотой ее появления, то есть относительной частотой, вдоль вертикальной оси. [8]
Этот компонент системы лишь неявно учитывался при рассмотрении конкуренции особей, а также выступал в виде жертвы в модели взаимодействующих популяций Вольтерра. [9]
Взаимная синхронизация колебаний играет большую роль в популяциях клеток, каждая из которых представляет собой отдельный автогенератор. Возможность регистрации колебаний в таких популяциях зависит от наличия частотной н фазовой синхронизации. Их затухание часто происходит вследствие рассинхронизации, а не действительного затухания в отдельных генераторах. Модель популяции, состоящая из двух типов генераторов, была рассмотрена Романовским и Черпавским ( 1971), которые дали оценку ширины полосы самосинхронизации в зависимости от коэффициента связи. Коэффициент связи в популяции клеток определяется проницаемостью мембран. [10]
Верхоф, так же как и Рамкришна с соавторами, не был последовательным на пути использования принципов формальной химической кинетики для моделирования роста популяции. Составив на основании феноменологии процесса схему предполагаемого перехода субстрата в биомассу, при выводе уравнения скорости роста эти исследователи фактически отказались от нее. Уравнения скорости составлены не в соответствии с постулатом: кинетики, а скомбинированы из известных для отдельных стадий; перехода уравнений и типа уравнения Моно. Это структурированная модель дискретной популяции, в которой в качестве основного акта рассматривается взаимодействие субстрата ( в обобщенном понимании) с активной субстанцией, представляющей собой своеобразный предшественник клеток. Результатом взаимодействия активной субстанции с субстратом является образование клеток, а в качестве регулирующего фактора роста всей популяции - сами клетки, отношение концентрации которых к оставшемуся субстрату, а также и их состояние определяют переход популяции из одной фазы роста в другую. [11]
Когда поколения перекрываются, RQ указывает лишь среднее число потомков, оставляемых одной особью, и ничего более. R - основная чистая скорость воспроизводства, - Уравнения, основанные на R и описывающие возрастание численности популяции с течением времени. Если зависимость плодовитости и выживания от возраста неизменна, то популяция достигает постоянной удельной скорости роста, равной г, и неизменного возрастного состава. Пример расчета обобщающих показателей применительно к популяции организмов с перекрывающимися поколениями, - Модель популяции, удельная скорость роста которой стремится к г, а возрастная структура - к стационарной. Почему когортные и статические таблицы выживания, как правило, не одинаковы. [12]
Страницы: 1