Математическая модель - рост - популяция
Cтраница 1
Математическая модель роста популяции, созданная в предположении независимости элементарных актов клеточного уровня ( бинарного деления) от взаимного влияния особей, не способна описать поведение популяции в реальных условиях ее существования. Переход к попыткам описания закономерностей именно на популяционном уровне неизбежно приводит к тому, чтобы отказавшись от идеализированной схемы, учитывать те закономерности, которые характеризуют этот уровень биологических систем. Такой качественной особенностью популяции как локального представителя вида является внутривидовая борьба, которая может принимать формы взаимного ингибирующего влияния, своеобразного каннибализма, конкуренции за субстрат; к тому же само по себе исчерпание субстрата при культивировании микроорганизмов периодическим способом также должно учитываться при анализе процессов роста и размножения микроорганизмов. [1]
Хотя приведенные выше математические модели роста популяции и учитывали качественную особенность популяционно-го уровня - внутривидовую борьбу, которая связывалась с тем или иным проявлением взаимодействия особей, тем не менее их все можно отнести только к моделям портретного типа. Безусловно, появление таких подходов следует рассматривать как определенный прогресс по сравнению с поисками эмпирических зависимостей или моделей типа уравнения экспоненциального роста популяции. Однако рассмотренные зависимости описывают только лишь накопление биомассы в предположении внут-рипопуляционного взаимодействия особей ( влияющих либо непосредственно на процессы размножения или гибели, либо опосредствованно через продукты метаболизма), но не вскрывают сущности процесса перехода субстрата в организационную биомассу популяции. Модели рассмотренного типа построены при неявном предположении протекания процесса роста популяции в условиях практически неограниченного количества субстрата. Вместе с тем очевидная роль питательной среды в процессах роста и размножения микроорганизмов безусловно должна быть отражена в соответствующих моделях развития популяций. [2]
Следует отметить, что математическая модель роста популяции, полученная для упрощенной схемы процесса ( XII), включает зависимость Xf ( M0, K, / Cs, 0 в явном виде; при этом смешанный параметр ftiM - 0, возникший в результате исключения из рассмотрения промежуточного этапа роста, имеет смысл обобщенной константы. [3]
Поэтому неслучайно при создании математических моделей роста популяции авторы очень часто обращались к кинетике ферментативных процессов, подбирая уже готовые выражения для зависимости скорости ферментативной реакции от содержания компонентов в исследуемой системе. При этом установленные строгим путем закономерности в бесструктурных системах переносились на протекание реакции в узком месте цепи внутриклеточного микробиологического синтеза без раскрытия особенностей механизма реакций, протекающих в узком месте. [4]
Нельзя сказать, что рассмотренные выше математические модели роста популяций совсем не относились к моделям кинетического типа. В том или ином виде все они включают выражение для скорости роста в виде дифференциального уравнения, интегральная форма которого описывает или должна описывать изменение во времени концентрации рассматриваемого компонента системы. [5]
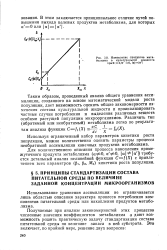
При оценке результатов экспериментальной проверки математической модели обратимого автокаталитического роста популяции, основные положения которой изложены в предыдущих разделах, целесообразно рассмотреть следующие зависимости: относительной скорости роста популяции от ее численности; относительной скорости образования микроорганизмов ( синтеза биомассы) от численности популяции; равновесной концентрации микроорганизмов от запаса субстрата в питательной среде; максимальной относительной скорости роста от запаса субстрата в питательной среде. [6]
Таким образом, для расширения области применения математической модели автокаталического обратимого равновесного роста популяции ( в частности, для описания начального периода роста) необходимо рассмотреть более подробную схему, учитывающую этапы преобразования субстрата питательной среды в промежуточный внутриклеточный субстрат, или, другими словами, перейти к рассмотрению кинетики сложных реакций. При этом может получить рациональное истолкование и кажущееся неподчинение процесса роста популяции закону действующих масс, что выражается в независимости общей скорости процесса от концентрации исходных компонентов и в общем-то является одним из отличительных признаков сложного процесса, проходящего через ряд промежуточных состояний. [7]
Уравнения ( XXXI) и ( XXXII) математической модели роста популяции, выведенные на основании совместного рассмотрения уравнения ассимиляции и уравнения сорбции ( десорбции) газообразных компонентов, дают возможность оценить требуемую величину объемного коэффициента массопередачи в зависимости от планируемой предельной численности популяции А, физиологической характеристики микроорганизма а, константы его скорости роста и заданного уровня содержания компонентов питательной среды или продуктов метаболизма. [8]
Однако, рассматривая вопросы, относящиеся к выбору математических моделей роста популяции, необходимо иметь в виду следующее положение. Изменения, происходящие и регистрируемые в биологических системах, являются результатами процессов, в которых участвуют компоненты различных уровней иерархической структурно-системной организации. Поэтому важным методологическим вопросом математического моделирования и является выбор уровня организации биосистемы, моделирование взаимодействия компонентов которого наиболее полно и адекватно соответствует поставленной при математическом моделировании практической задаче. [9]
Расчет такого режима легко проводится по уравнению ( XVII) математической модели роста популяции, а осуществление этого режима можно проводить на всех этапах технологической разработки условий культивирования. [10]
 |
Накопление продуктов метаболизма в культуральной жидкости прио о а 0, а о. [11] |
Таким образом, проведенный анализ общего уравнения ассимиляции, созданного на основе математической модели роста популяции, дает возможность оценить общие закономерности изменения состава культуральной жидкости и проанализировать частные случаи потребления и выделения различных веществ особями растущей популяции микроорганизмов. [12]
Вместе с тем было установлено, что некоторые наблюдаемые закономерности роста не следуют из структуры математической модели автокаталитического обратимого роста популяции. Так, при заданном уровне схематизации модели уравнение роста не описывает лаг-фазу и справедливо лишь с момента времени культивирования, когда начинается регулярный рост биомассы. Кроме того, согласно рассматриваемой математической модели, должна наблюдаться прямая связь между запасом субстрата в питательной среде и максимальной удельной скоростью роста популяции, однако на практике этого не наблюдается: независимо от содержания субстрата в питательной среде М0 параметр PI. MO сохраняет постоянное значение для данного зида микроорганизма и данного качественного состава питательной среды. [13]
В настоящей главе рассматриваются некоторые примеры моделей реального мира, иллюстрирующие теорию, развитую в предыдущих главах. В параграфе 5.1 описываются математические модели роста конкурирующих популяций, а также модели хищник - жертва и исследуются свойства устойчивости ненулевого равновесия. В параграфе 5.2 изучаются дискретные модели, возникающие в экономике, и анализируется экспоненциальная устойчивость состояния равновесия, отвечающего работающей системе. В параграфе 5.3 рассматриваются консервативные механические системы с некоторыми диссипативнымк силами и изучаются свойства их устойчивости. В параграфе 5.4 представлены модели, взятые из экономики, и с помощью метода вектор-функций Ляпунова доказывается, что рынок стремится к некоторой заданной эволюции независимо от начальных условий. В параграф 5.5. анализируются упругопластичные модели, которые приводят t линейной интегродифференциалыюй системе, и изучается ее устой чивость путем использования метода приведения этой системы к экви валентной линейной дифференциальной системе. Параграф 5.6 посвя щен моделям химической кинетики. Здесь для исследования асимпто тической устойчивости решений реактивно-диффузионной системь применяется метод сравнения с функционалом Ляпунова. В заключи тельном параграфе 5.7 рассматриваются линейная и нелинейная мо дели вооружений и разоружения. [14]
 |
Установка НКМ-2 для непрерывного культивирования клеток млекопитающих. [15] |
Страницы: 1 2