Молекула - колицин
Cтраница 1
Молекулы колицина присоединяются к специальным рецепторам - молекулам липополисахаридам, выступающим из внешней жесткой оболочки бактерии. Присоединив молекулу колицина К ( или Е1), бактерия некоторое время продолжает функционировать нормально. [1]
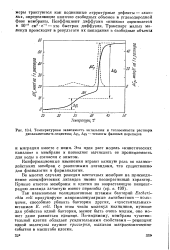 |
Температурная зависимость энтальпии и теплоемкости раствора, дипальмитоил-сс-лецитина. Д. 1, Д. 2 - теплоты фазовых переходов. [2] |
При этом число молекул колицинов, нужное: для убийства одной бактерии, может быть очень малым, оно может даже равняться единице. По-видимому, мембрана чувствительной клетки обладает усилительными свойствами - рецепцияс одной молекулы служит триггером, вызывая макроскопические-события в масштабе клетки. [3]
Для объяснения наблюдаемых эффектов влияния молекул колицина на нормальные и мутантные бактерии E-coli делается предположение, что внедрение молекулы колицина в клетку сопровождается снижением мембранной разности потенциалов. Снижение разности потенциалов приводит к прекращению активного транспорта лактозы, глютамина, ионов калия и магния через мембрану. В нормальной бактериальной клетке фермент АТФ-аза, находящийся в мембране в безнадежной попытке восстановить необходимую разность потенциалов, производит интенсивный гидролиз молекул АТФ, снижая количество этих молекул внутри клетки. Следствием уменьшения числа молекул АТФ является прекращение синтеза молекул белка, ДНК и РНК. [4]
Вопрос о молекулярном механизме происходящих в мембране изменений при присоединении молекул колицина остается открытым. Возможно, что снижение разности потенциалов обусловлено конформационной перестройкой белковых молекул, участвующих в активном транспорте. [5]
В мутантной клетке, из-за отсутствия фермента АТФ-азы при присоединении молекулы колицина, гидролиз молекул АТФ в мембране не происходит. [6]
Бактерия теряет способность к активному транспорту лактозы и глютамина ( рис. 32), когда молекула колицина проникает через цитоплазматическую мембрану. Одновременно внутри клетки резко уменьшается число молекул АТФ и прекращается синтез белков и молекул ДНК и РНК. [7]
Для объяснения наблюдаемых эффектов влияния молекул колицина на нормальные и мутантные бактерии E-coli делается предположение, что внедрение молекулы колицина в клетку сопровождается снижением мембранной разности потенциалов. Снижение разности потенциалов приводит к прекращению активного транспорта лактозы, глютамина, ионов калия и магния через мембрану. В нормальной бактериальной клетке фермент АТФ-аза, находящийся в мембране в безнадежной попытке восстановить необходимую разность потенциалов, производит интенсивный гидролиз молекул АТФ, снижая количество этих молекул внутри клетки. Следствием уменьшения числа молекул АТФ является прекращение синтеза молекул белка, ДНК и РНК. [8]
Опыты с мутантными бактериями E-coli, лишенными фермента АТФ-азы в цитоплазматической мембране, показали, что при присоединении к ним молекул колицина их поведение существенно отличается от поведения обычной бактериальной клетки. После присоединения молекулы колицина к таким мутантным клеткам также происходит прекращение активного транспорта лактозы, глютамина и ионов К и Mg2, однако внутри клетки не прекращается синтез белков и молекул ДНК и РНК. При этом количество молекул АТФ внутри клетки даже возрастает. [9]
Молекулы колицина присоединяются к специальным рецепторам - молекулам липополисахаридам, выступающим из внешней жесткой оболочки бактерии. Присоединив молекулу колицина К ( или Е1), бактерия некоторое время продолжает функционировать нормально. [10]
Опыты с мутантными бактериями E-coli, лишенными фермента АТФ-азы в цитоплазматической мембране, показали, что при присоединении к ним молекул колицина их поведение существенно отличается от поведения обычной бактериальной клетки. После присоединения молекулы колицина к таким мутантным клеткам также происходит прекращение активного транспорта лактозы, глютамина и ионов К и Mg2, однако внутри клетки не прекращается синтез белков и молекул ДНК и РНК. При этом количество молекул АТФ внутри клетки даже возрастает. [11]
Переход от активного состояния клетки бактерии ко второму, неактивному, состоянию происходит только в том случае, когда цнтоплазматическая мембрана находится в энергизированном состоянии. Следовательно, для прохождения молекулы колицина через мембрану требуется затрата энергии. Скорость перехода зависит от состояния липидов в мембране. Переход затормаживается при охлаждении мембраны или включении в нее липидов с насыщенными углеродными связями, которые делают липидный слой более жестким. Цитоплазматическая мембрана бактерии переходит в энергизированное состояние за счет энергии двух независимых процессов: окисления молочной и янтарной кислот ( перенос электрона к кислороду) и гидролиза молекул АТФ. Энергизация мембраны за счет энергии гидролиза осуществляется специальным ферментом АТФ-азой, находящимся в мембране. Этот же фермент может осуществлять обратную реакцию - синтез молекул АТФ из молекул АДФ и фосфата за счет энергии мембраны. [12]
Страницы: 1