Развернутая молекула
Cтраница 1
 |
Модельное изображение процесса ориентации и релаксации фибриллярной структуры полигексаметиленадипинамида. [1] |
Развернутые молекулы агрегируются в пачки взаимноориентиро-ванных цепей, которые в дальнейшем, при образовании более сложных структур, играют роль независимых структурных единиц. Следовательно, при образовании более сложных структур роль структурных единиц выполняют глобулы или пачки цепей. [2]
Но сильное взаимодействие развернутых молекул с поверхностью в начале формирования поверхностной пленки, ограничивая подвижность цепей, препятствует формированию на поверхности надмолекулярных структур, и сами они и их упаковка на поверхности могут быть более рыхлыми. [3]
Для этого к цепи развернутой молекулы ДНК пристраивается в строгом порядке, определяемом последовательностью оснований ДНК, цепь РНК - посредника. Так наследственная информация считывается с ДНК. Теперь уже начинается сам процесс биосинтеза белка: другой вид РНК - - транспортная РНК, характерная для каждой аминокислоты и образующая с ней ковалентную связь, ставит аминокислоту в нужное место пептидной цепи в соответствии с инструкцией, полученной от ДНК через РНК - посредника. Само собой разумеется, что синтез белка идет при участии ферментов. [4]
 |
Схематическое изображение белковых молекул на поверхности раздела.| П-40. Полипептидный монослой в виде це -.,. [5] |
При очень низких поверхностных давлениях в белковой пленке вполне могут существовать развернутые молекулы, и Зингер [171] считает, что в таких условиях применима теория Флори - Хаггинса для полимерных растворов. При плотной упаковке один сегмент цепи занимает площадь Ъ, средняя площадь пленки, приходящаяся а сегмент, равна о. Длинная гибкая цель, предоставленная самой себе, не является ни полностью развернутой, ни полностью свернутой. Она находится в некотором промежуточном состоянии, скорее всего почти развернутом. При сжатии цепь становится более компактной; при этом конфигурационная энтропия уменьшается. Таким образом, как п, так и z должны зависеть от давления. [6]
ОКЭМ, характерны ассоциаты анизодиаметричного типа, образованные, по-видимому, из развернутых молекул. Для олигокарбонатметакрилата с гибкими блоками ОКДМ наблюдаются глобулярные структуры, возникновение которых связано со сворачиванием пучков молекул. [7]
Если, например, полностью развернуть молекулу рибонуклеа-зы путем восстановления четырех ее дисульфидных мостиков меркаптоэта-нолом в 8 М мочевине, а затем вызвать реокисление таких развернутых молекул в контролируемых условиях, то молекулы ( от 95 до 100 %) вновь приобретают нативную конформацию, что подтверждается восстановлением не только физических свойств, но и ферментативной активности. Этот опыт схематически представлен на фиг. В табл. 20 приведены данные по рена-турации некоторых белков. Во всех случаях, за исключением инсулина, степень восстановления нативных структур значительно превышает величину, которой следует ожидать, исходя из статистических соображений. Эти данные вовсе не означают, однако, что процесс образования дисульфидных связей в белках может протекать in vivo без направленного катализа. [8]
В работах [1-4] при исследовании строения полимеров было показано, что существуют два типа элементарных структурных единиц, которые являются основой для создания более сложных структур: глобулы, представляющие собой свернутые молекулярные цепи, и пачки, образующиеся при агрегации развернутых молекул. Уже сравнительно хорошо изучены условия образования кристаллов высокомолекулярных соединений на ряде полимерных веществ. В то же время известно, что в зависимости от условий приготовления и природы полимера образуются самые разнообразные вторичные структуры. Изучение вторичных структур полимеров обычно проводилось путем приготовления образцов из разбавленных растворов. [9]
Вот теперь, наконец, вступают в действие антикодоны. К кодонам развернутой молекулы мРНК пристраиваются не сами Аминокислоты, а комплементарные антикодоны. Эти последние не существуют как таковые совершенно независимо, но входят в состав молекулы РНК третьей и пока последней известной разновидности. Исходя из ее функции, эту РНК назвали транспортной ( сокращенно тРНК); в качестве синонимов употребляют также названия растворимая РНК и РНК-переносчик. Молекулы транспортной РНК сначала выбирают из фонда аминокислот, производимых клеткой, нужную аминокислоту и затем, нагруженные каждая своей аминокислотой, направляются к рибосомам. Итак, имея в своем составе определенный антикодон, данная молекула тРНК находит свое место на молекуле мРНК; сюда она и доставляет соответствующую аминокислоту. Молекулы транспортной РНК состоят примерно из 80 нуклеотидов, соединенных в цепочку. Аминокислота присоединяется к тому концу молекулы тРНК, на котором стоит триплет ЦЦА. [10]
Наличие глобулярных структур уменьшает возможность контакта молекулы с поверхностью адсорбента, что сильно снижает прочность возможной связи с поверхностью и приводит к предпочтительной сорбции растворителя. Действительно, для более развернутых молекул желатины адсорбция наблюдается. Очевидно, аналогично можно объяснить и данные по адсорбции сополимеров стирола с метакриловой кислотой. Сополимер с большим содержанием карбоксильных групп, вероятно, образует в растворе достаточно устойчивые глобулярные структуры за счет внутримолекулярного взаимодействия, что приводит к отсутствию адсорбции. Для сополимера с небольшим числом СООН-групп адсорбция идет из хорошего растворителя, но не идет из плохого. [11]
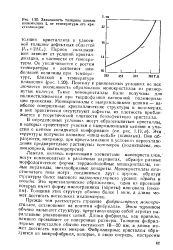 |
Зависимость толщины ламели полиэтилена Ь от температуры его кристаллизации. [12] |
Период складывания зависит от условии кристаллизации, в частности от температуры. Поэтому к равновесных условиях не иск-лючается возможность образования монокристалла из развернутых молекул. Такие монокристаллы были получены для полимстнлсноксида при твердофазной катнонной полимеризации триоксана. Монокристаллы с выпрямленными цепями - наиболее совершенные кристаллические структуры в полимерах, В них практически отсутствуют дефекты, их плотность приближается к теоретической плотности бездефектного кристалла. [13]
Эта область неполных, но достаточно стабильных равновесий полимерных студней очень важна, так как часто можно получить из одного и того же полимера растворы одинаковой концентрации, годные или негодные для определенных технических целей, в частности, для применения в качестве клеев или прядильных растйоров. Понимание этой интересной области явлений позволяет справиться с практически нежелательными явлениями, а также раскрывает некоторые стороны биологических процессов, в которых цепные макромолекулы, свернутые в глобулы, проявляют необходимую подвижность, несвойственную развернутым молекулам. [14]
На основании этих фактов укрепилось мнение [1] о том, что оболочка жирового шарика имеет следующую структуру. Первичный слой состоит из монослоя радиально ориентированных молекул фосфолипидов с углеводородными радикалами в жировой фазе и гидрофильными группами в водной. Затем следует слой из развернутых молекул оболочечного протеина, цепи которого соприкасаются в некоторых местах с поверхностью жирового шарика. Цепи полипептида свернуты и скручены различным образом, но не исключено, что в некоторых местах цепи располагаются упорядочение. Ионизированные группы могут образовывать связи с фос-фолипидами благодаря электростатиче-ским силам. [15]
Страницы: 1 2