Живая мышца
Cтраница 1
Живая мышца давно служит заманчивым прототипом для построения полимерных преобразователей, которые приближались бы к ней по принципу действия. [1]
В живой мышце это взаимодействие контрактильного белка с АТФ, очевидно, становится возможным лишь в момент передачи нервного возбуждения с концевых моторных пластинок на мышечное волокно. Что происходит в мышце в этот момент и почему имеющаяся в невозбужденной мышце АТФ не вызывает сокращения контрактильного белка, остается еще не вполне ясным. [2]
В живой мышце органом управления является нервная система. К настоящему времени в лабораторных условиях создан полимер, обладающий свойством ионной проводимости в осевом направлении. [3]
В живой мышце существует весьма тонкий механизм, питающий ее горючим материалом и выделяющий продукты сгорания. Для синтетической мышцы такой механизм находится еще в стадии разработки. [4]
В живой мышце это взаимодействие контрактильного белка с АТФ очевидно, становится возможным лишь в момент передачи нервного возбуждения с концевых бляшек на мышечное волокно. Что происходит в мышце-в этот момент и почему имеющаяся в невозбужденной мышце АТФ не вызывает сокращения контрактильного белка, остается еще не вполне ясным. [5]
Возможность пребывания живой мышцы в расслабленном состоянии при наличии в ней достаточно высокой концентрации АТФ объясняется снижением в результате действия кальциевой помпы концентрации ионов Са2 в среде, окружающей миофибриллы, ниже того предела, при котором еще возможны проявление АТФазной активности и сократимость акто-миозиновых структур волокна. Быстрое сокращение мышечного волокна при его раздражении от нерва ( или электрическим током) является результатом внезапного изменения проницаемости мембран и как следствие выхода из цистерн и трубочек саркоплазматической сети и Т - системы некоторого количества ионов Са2 в саркоплазму. [6]
Прежде всего, следует отметить следующую особенность живой мышцы. [7]
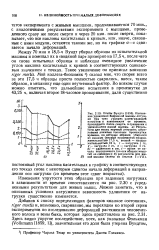
Изменения упругости этих образцов при заданных нагрузках в зависимости от времени сопоставляются на рис. 2.30 с усредненными результатами для живых мышц. Можно заметить, что в описанных условиях нагружения зависимость удлинения от нагрузки существенно изменяется. [9]
Однако представление о возможности использования при сокращении мышцы энергии АТФ в процессе Г - Ф - превращения актина не получило подтверждения в работах американского биохимика Джерджели, по данным которого в живой мышце актин находится в Ф - форме. В связи с этим ряд авторов ( Оосава и др.) пытаются заменить представление Штрауба концепцией о возможности линейно-спиральной трансформации полимерного актина в сократительном акте. Эта теория построена на новых данных о структуре Ф - актина. Как оказалось, Ф - актин представляет собой двухспиральную структуру, каждая цепочка которой состоит из множества бусинок - глобул мономера Г - актина. Цепочки могут существовать как в спиральной форме, так и вытянутой - линейной. По данным Оосава, изменение конфигурации и длины двухспиральной цепочки Ф - актина связано с разрывом части мостиков между мономерами актина соседних цепочек и происходит при взаимодействии с АТФ, который при этом, по-видимому, подвергается расщеплению. [10]
Ввиду исключения реакций окислительного распада исключаются реакции ресинтеза гликогена и органических фосфорных соединений и уже в первые часы накапливается значительное количество молочной кислоты и ортофосфатов, что, в свою очередь, приводит к резкому изменению нейтральной реакции живой мышцы ( близкой к рН 7 0 - 7 2) в кислую сторону ( рН 6 0 - 6 2), большему, чем то, которое наблюдается при утомлении живой мышцы. Одновременно с этим наблюдается переход значительной части фракции солерастворимых белков в нерастворимое состояние. При этом происходит дальнейшее уменьшение содержания гликогена и прогрессивное накопление неорганического фосфата и редуцирующих Сахаров. Содержание молочной кислоты, хотя и медленно, нарастает. Одновременно происходит переход части нерастворимых белков в растворимое состояние, однако сколько-нибудь глубокого распада белков не наблюдается. Резкое же увеличение содержания аммиачного азота свидетельствует о начавшихся процессах микробной порчи мяса. [11]
Ввиду исключения реакций окислительного распада исключаются реакции ресинтеза гликогена и органических фосфорных соединений и уже в первые часы накапливается значительное количество молочной кислоты и ортофосфатов, что, в свою очередь, приводит к резкому изменению нейтральной реакции живой мышцы ( близкой к рН 7 0 - 7 2) в кислую сторону ( рН 6 0 - 6 2), большему, чем то, которое наблюдается при утомлении живой мышцы. Одновременно с этим наблюдается переход значительной части фракции солерастворимых белков в нерастворимое состояние. При этом происходит дальнейшее уменьшение содержания гликогена и прогрессивное накопление неорганического фосфата и редуцирующих Сахаров. Содержание молочной кислоты, хотя и медленно, нарастает. Одновременно происходит переход части нерастворимых белков в растворимое состояние, однако сколько-нибудь глубокого распада белков не наблюдается. Резкое же увеличение содержания аммиачного азота свидетельствует о начавшихся процессах микробной порчи мяса. [12]
Активность фактора Марша - Бендалла, изменяющего аденозинтри-фосфатазную активность миозина, в свою очередь зависит от ряда уело -, вий, в частности от концентрации ионов Mg и Са в мышечных волокнах. Концентрация же этих ионов, особенно Са - ингибитора фактора Марша, в, живой мышце может чрезвычайно быстро изменяться под влиянием нервных импульсов. [13]
Мы обсуждаем здесь работу Вундта не только в связи с замечаниями Фолькмана по поводу несправедливых нападок Вундта на Вертгейма, но и потому, что два качественных аспекта его экспериментов обсуждаются иногда неудачно. Одним из них является его сравнение зависимостей между напряжениями и деформациями для мышц бедра живой лягушки и для такой же мышцы, испытанной в промежутке 30 мин после смерти лягушки, с учетом влияния как усталости, так и постоянства нагрузки на живую мышцу. Второй интересный аспект экспериментов Вундта состоит в его исследовании, на образцах из человеческого волоса, а также вены быка, зависимости между первичными и вторичными деформациями, с учетом упругого последействия, в условиях наличия последовательности многочисленных приращений при нагружении и разгрузке. [14]
Остальные мышечные белки находятся в незначительных количествах. Приведенное распределение основных фракций мышечных белков не является постоянным и значительно меняется в процессах посмертных изменений мышечной ткани, сопровождающихся переходом части растворимых белков в нерастворимое состояние. В живой мышце миоген и глобулин X входят в состав саркоплазмы, а миозин и актин - в состав мио-фибриллей. [15]
Страницы: 1 2