Жакоба
Cтраница 2
Регуляция транскрипции далеко не всегда реализуется по схеме Жакоба и Моно посредством репрессора, негативного ре-гуляторного фактора. [16]
Следующим важным шагом в изучении информационной РНК явился опыт Бреннера, Жакоба и Месельсона, о котором рассказано выше. Было показано, что рибосомы - универсальный неспецифический аппарат для синтеза белков, причем информационная РНК, образуясь, по-видимому, в хромосоме, устремляется из ядра & цитоплазму, прикрепляется к рибосомам и направляет их действие в сторону синтеза определенных специфических белков. [17]
К настоящему времени сравнительно хорошо аргументирован молекулярный механизм генной регуляции у бактерий. Жакоба и Ж - Моно [8], структурные гены, работающие на синтез белка, объединены в группы или опероны. [18]
Таким образом, этот фактор может существовать как в интегрированном, так и в полуавтономном состоянии. По предложению Жакоба, Вольмана и Моно подобные генетические элементы были названы эписомами. [19]
Подробное исследование симметричных триггеров Жакоба - Моно было предпринято Черна век им, Григоровым и Поляковой ( 1967), которые указали на большую роль этих механизмов в процессе дифферен-цировки тканей. Последние являются значительно более эффективными регуляторами. [20]
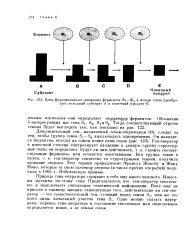 |
Цепь функционально связанных ферментов Ii - Ф4 в четыре этапа преобразует исходный субстрат А в конечный продукт Е. [21] |
Дополнительный ген, называемый геном-оператором ( О), следит за тем, чтобы группа генов Sj - S4 выключалась одновременно. Этот термин принадлежит Франсуа Жакобу и Жаку Моно, которые за свою гипотезу оперона ( в числе прочих открытий) получили в 1965 г. Нобелевскую премию. [22]
Пусть для этой последовательности нужно три фермента. Тогда по схеме Моно и Жакоба этот продукт вступает в реакцию с одним из белков, производимым на особом участке ДНК, называемом геном-регулятором. Это соединение, так называемый репрессорный белок, в свою очередь, действует на систему, состоящую из участка ДНК, называемого оператором ( ген-оператор), и участков, называемых структурными генами. На структурных генах и получается м - РНК, непосредственно используемая для синтеза требуемых белков-ферментов. Роль гена-оператора заключается в контролировании скорости использования структурных генов для синтеза белка. Если репрессор соединился с геном-оператором, блокировал его, то работа соответствующих структурных генов прекращается. В нашем примере ген-оператор должен контролировать деятельность трех участков ДНК, производящих нужные м - РНК и, следовательно, и ферменты для данной сложной реакции. Выключение оператора в результате фиксации на нем белка репрессора, связанного с продуктом реакции Р, прекращает я производство ферментов для реакции. Если концентрация продукта Р понизилась, оператор освободился от блокады, то синтез ферментов возобновляется и вся биохимическая машина запускается вновь. [23]
За последние годы получены новые экспериментальные материалы, которые в некоторой степени проливают свет на эту сложную проблему - регуляцию синтеза белка на клеточном уровне. Эти достижения связаны прежде всего с исследовательской группой Моно и Жакоба, работающей в Пастеровском институте. В регуляции белкового синтеза на клеточном уровне нуклеиновые кислоты, видимо, также играют решающую роль. Прежде всего, было найдено, что различные участки молекулы ДНК функционально неоднородны и одна молекула ДНК может определять синтез большого числа функционально и химически различных белков клетки. [24]
В настоящее, время считают, что регуляции подвержены все или почти все этапы биосинтеза белков. Например, метаболиты и гормоны могут изменять сродство белков-репрессоров к регуляторным отделам ДНК; гормоны способны модифицировать активность метилаз, участвующих в биосинтезе рРНК; новообразованные белки способны активировать рибонукле-азы и тем самым ускорять распад своих мРНК и т.п. Согласно теории Жакоба и Моно, в биосинтезе белков у бактерий участвуют три типа генов: структурные гены, ген-оператор и ген-регулятор. Структурные гены определяют первичную структуру белков. Формирование мРНК начинается с промотора и далее распространяется вдоль оператора и контролируемых им структурных генов. Оператор и структурные гены называют опероном. Деятельность оперона контролируется геном-регулятором. Оперон и ген-регулятор находятся в разных участках цепи ДНК, поэтому связь между ними осуществляется с помощью белка-репрессора, синтезируемого по информации гена-регулятора. Если репрессор связан с геном-оператором, то РНК-по-лимераза не может синтезировать мРНК, а следовательно, не синтезируются и белки. Если ген-оператор свободен, процесс транскрипции возможен и информация структурных генов используется для синтеза белков. [25]
Было известно, что синтез нового белка в клетке, например, после индукции фермента или после заражения фагом, начинается через время порядка минуты. Так, после скрещивания бактерий HfrLac X F-Lac - и уничтожения мужских клеток обнаруживался синтез фермента 5-галактозидазы в женских клетках, получивших ген Lac, через несколько минут после конъюгации. Это и заставило Парди, Жакоба и Моно 188 впервые выдвинуть идею о нестабильном промежуточном продукте, служащем матрицей для синтеза белка. [26]
Ну а что же, эффект отрицательной обратной связи присущ только целостным организмам. Нет, по его принципу работают все клетки, начиная с вирусов и бактерий и кончая клетками человеческого тела. В 1965 г. французским биохимикам Франсуа Жакобу и Жаку Моно была присуждена Нобелевская премия за разработку теории клеточной репрессии. Суть ее состоит в следующем. Клетка сама себя восстанавливает из белковых продуктов, которые она строит из отдельных аминокислот, набор которых ограничен и достаточно стереотипен. Специфика построения клеточного белка определяется тремя типами генов: структурным геном, в котором хранится шифр специфичности, геном-оператором, запускающим процесс, и геном-репрессором, останавливающим процесс наработки белка, когда в нем отпадает необходимость. [27]
Таким образом, найдя колонию рекомбинантов на чашке Петри, мы узнаем, из какой колонии исходных клеток F происходят клетки, приведшие к рекомбинантам. Аналогичным образом, выбирая другой ауксотрофный мутант в качестве женской культуры, мы отбираем из той же популяции F 1 другие мутанты Hfr с началом хромосомы вблизи локусов Lac, Тгур, Вь М и др. Тем самым основные положения гипотезы Воль-мана и Жакоба оказываются доказанными. [28]
Само предположение о необхфгямости индуктора для синтеза белка так же хорошо соответствует фактам, как предположение о наличии репрессора в диком типе клеток. В недавней работе Парди, Жакоба и Моно этот вопрос подвергнут анализу. Чтобы понять руководящую идею их работы, необходимо напомнить одно из важных понятий генетики - понятие доминантности признака. [29]
Тогда можно ожидать, что в любой популяции клеток F присутствуют в качестве небольшой примеси разнообразные мутанты Hfr, у которых начало хромосомы О совпадает с любыми маркерами. Подобная гипотеза объясняет сразу многое. Во-первых, понятно, что Ледерберг, работая с культурой F как донорной, мог измерить расстояния на генной карте по рекомбинации маркеров. Из гипотезы Вольмана и Жакоба был выведен ряд следствий, которые можно было подвергнуть экспериментальной проверке. [30]
Страницы: 1 2