Паращитовидная железа
Cтраница 1
Паращитовидная железа считается наиболее радиорезистентной из всех органов внутренней секреции. В литературе имеется крайне мало-сообщений о действии ионизирующего излучения на эту железу. [1]
Опухоли паращитовидной железы после общего или местного облучения не развиваются. [2]
В паращитовидной железе синтезируются два полипептидных гормона - паратиреоидный ( парат) гормон и кальцитонин. Местом синтеза кальцитони-на является также щитовидная железа. [3]
Паратиреоидный гормон паращитовидной железы наряду с кальцитонином и 1 25-дигидроксихолекальциферолом участвует в регуляции фосфатного и кальциевого обмена у млекопитающих. Снижение содержания кальция в крови вызывает секрецию паратиреоидного гормона паращитовидной железой. Это повышает содержание кальция в крови и стимулирует секрецию фосфатов в мочу, чем снижается их содержание в крови. [4]
Паратгормон был получен в частично очищенном состоянии в 1925 г. из паращитовидной железы быка, и только полвека спустя его получили в гомогенном состоянии. Он представляет собой полипептид, состоящий из 84 аминокислотных остатков, причем биологическая активность определяется TV-концевым фрагментом, состоящим из 34 аминокислот. [5]
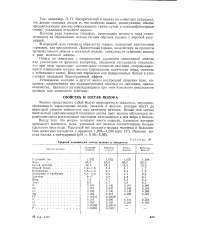 |
Средний химический состав молока в процентах. [6] |
Установлено влияние и других желез внутренней секреции ( как, например, надпочечника или паращитовидной железы) на лактацию, однако неизвестно, являются ли наблюдающиеся при этом изменения результатом прямого или косвенного действия. [7]
Состояние гипокаль-циемии наблюдается при авитаминозе D ( рахите), у беременных, при недостаточной функции паращитовидной железы, заболеваниях почек, при отравлениях фторидами. [8]
К синдрому Ди Джорджи следует относить [ Carey, 1980 ] только случаи с комплексом поражений тимуса, сердечно-сосудистой системы и паращитовидной железы. Автор описал различные варианты этого синдрома, наследуемые аутосомно-доминантно, аутосомно-рецессивно, а также синдромы Ди Джорджи, сопровождаемые хромосомными аномалиями. [9]
Щитовидная и молочная железы, хотя они и легко доступны ультразвуковому обследованию, часто требуют использования водяного и иного буфера, чтобы на изображение не повлияли аномалии ближней зоны поля. При исследовании щитовидной и ( реже) паращитовидной железы основное применение ультразвука - различение кистозных и твердых образований, что возможно при хорошем подавлении шума и артефактов, вызванных реверберацией и боковыми лепестками излучателя. [10]
Паратиреоидный гормон паращитовидной железы наряду с кальцитонином и 1 25-дигидроксихолекальциферолом участвует в регуляции фосфатного и кальциевого обмена у млекопитающих. Снижение содержания кальция в крови вызывает секрецию паратиреоидного гормона паращитовидной железой. Это повышает содержание кальция в крови и стимулирует секрецию фосфатов в мочу, чем снижается их содержание в крови. [11]
АТФ в крови и нарушению процессов фос-форилирования; одновременно в 10 - 20 раз увеличивается содержание А. Предполагают, что в механизме нарушения фосфорно-кальциевого обмена важную роль играет гормон паращитовидной железы, повышающий всасывание А. Обнаружены конкурентные отношения между А. [12]
Паратиреоидный гормон синтезируется в виде полипептида - предшественника, состоящего из 115 аминокислотных остатков. В результате локального протеолиза отщепляется 31 аминокислотный остаток с N-копца и образуется активный гормон. Фактором, регулирующим содержание активного гормона в крови, является концентрация кальция и содержание пропаратгормона в клетках паращитовидной железы. В физиологических условиях большая часть пропаратгормона распадается в клетках, однако дефицит кальция приводит к уменьшению его распада и увеличению выхода активного гормона. Вновь образованный паратгормон поступает в секреторные гранулы и перемещается из клеток в кровь. Скорость секреции обратно пропорциональна концентрации кальция в плазме крови. Кроме того, на скорость освобождения гормона влияет уровень цАМФ в клетках паращитовидной железы. [13]
Кальцитонин является антагонистом паратгор-мона и ингибирует резорбцию костной ткани. Его биологическое действие реализуется по мембрано-опосредованному механизму и вызывает уменьшение концентрации кальция в плазме крови. КТ действует не только на минеральную составляющую костей, но и на их органический матрикс. Это проявляется в ингибировании костного коллагена, инактивации кислой фосфатазы и р-глюкуронидазы, а также активации щелочной фосфатазы. Кальцитонин способствует транспорту фосфора из крови в костную ткань для образования гидроксиаппатита в последней, а также оказывает выраженное действие на почки, подавляя канальциевую реабсорбцию кальция и фосфора. Биологическое действие гормонов паращитовидной железы проявляется на фоне действия на обмен кальция и фосфора таких гормонов, как глюкокортикоиды и со-матотропин. [14]
Паратиреоидный гормон синтезируется в виде полипептида - предшественника, состоящего из 115 аминокислотных остатков. В результате локального протеолиза отщепляется 31 аминокислотный остаток с N-копца и образуется активный гормон. Фактором, регулирующим содержание активного гормона в крови, является концентрация кальция и содержание пропаратгормона в клетках паращитовидной железы. В физиологических условиях большая часть пропаратгормона распадается в клетках, однако дефицит кальция приводит к уменьшению его распада и увеличению выхода активного гормона. Вновь образованный паратгормон поступает в секреторные гранулы и перемещается из клеток в кровь. Скорость секреции обратно пропорциональна концентрации кальция в плазме крови. Кроме того, на скорость освобождения гормона влияет уровень цАМФ в клетках паращитовидной железы. [15]
Страницы: 1