Медьсодержащий белок
Cтраница 1
Медьсодержащий белок, который для живых организмов является важным металлсодержащим ферментом, аналогом железосодержащих белков, участвует в переносе электронов, транспорте кислорода, окислите. [1]
Гемоцианин - это медьсодержащий белок, который присоединяет одну молекулу О2 на каждые два атома меди ( гл. Деокси-Си ( 1) - форма заметно не поглощает в видимой области. Таким образом, спектральные данные для оксигемоцианина полностью согласуются с моделью связывания О2 по механизму окислительного присоединения типа гемэритрина. [2]
Дофамингидроксилаза млекопитающих представляет собой медьсодержащий белок, косубстратом которого может служить только аскорбиновая кислота. [3]
Далее следует отметить наличие еще примерно 10 молекул пластоцианина - медьсодержащего белка ( 2 атома Си на молекулу; мол. К) и бензохинонов ( разнообразные пластохиноны; см. гл. XV), а также НАДФ и флавины ( см. гл. К системе I относятся, кроме того, каротиноиды. Поглощаемая ими световая энергия с высокой эффективностью передается хлорофиллу. Система I, функционирующая при наличии окисляющегося субстрата ДН2, сама по себе, без системы II, осуществляет реакцию фотовосстановления ( фоторедукция) подобно тому, как это происходит у водорослей, нуждающихся в доноре водорода. [4]
Уриказа - фермент, ответственный за это каталитическое превращение - представляет собой медьсодержащий белок ( см. гл. [5]
У многих низших животных, таких, как крабы и улитки, переносчиком кислорода является медьсодержащий белок гемоцианин. [6]
При другой наследственной патологии - болезни Вильсона, помимо общей гипераминоацидурии, отмечается снижение концентрации медьсодержащего белка - церулоплазмина - в сыворотке крови и отложение меди в мозге, печени, почках. Генетический дефект связан с нарушением синтеза церулоплазмина. Возможно, свободная медь образует комплексы с аминокислотами, которые не всасываются в почечных канальцах. [7]
При другом наследственном пороке обмена-гепатоцеребралъной дистрофии ( болезнь Вильсона), помимо генерализованной ( общей) гипер-аминоацидурии, отмечаются снижение концентрации медьсодержащего белка церулоплазмина в сыворотке крови и отложение меди в мозге, печени, почках. Генетический дефект связан с нарушением синтеза церулоплазмина. Возможно образование комплексов меди с аминокислотами, которые не всасываются в канальцах. Аналогичная гипераминоацидурия наблюдается при галактоземии, синдроме Лоу и других наследственных заболеваниях. Пониженная экскреция аминокислот описана при квашиоркоре. [8]
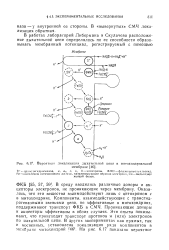 |
Вероятная локализация дыхательной цепи в митохондриальной. [9] |
ГГ - трансдегидрогеназа, а, ал, с, с, b - цптохромы, ФМН - флазипмоиопуклеотид, Fe - комплексы негечинового железа, катализирующие перенос электрона, Си - медьсодержащий белок. [10]
 |
Взаимосвязь некоторых фотосинтетических электронтранспортных цепей через большой фонд пластохинона ( PQ. Цит - цитохром. ПЦ - пла. [11] |
ЫАОР - оксидоредуктаза окрашен в желтый цвет, поскольку и качестве простетической группы содержит рибофлавин ( гл. Пластоцианин представляет собой медьсодержащий белок, и его интенсивная синяя окраска обусловлена, вероятно, поглощением в результате переноса заряда медью. Неокрашенный важный компонент системы - пластохинон (10.8) - представляет собой бензохинон с длинной изопреноидной боковой цепью ( свойства бензохинонов были описаны в гл. [12]
Далее в цепи переноса электронов от фотосистемы 11 к фотосистеме 1 принимают участие два цито. Следующим компонентом цепи является медьсодержащий белок пластоцианин, от которого электрон поступает в реакционный центр фотосистемы I. Комплекс, образованный нластохиноиом, цитохромами 6559 и С552 и пластоцнанином, по-видимому, является близким аналогом комплекса III в цепи переноса электронов в митохондриях. При прохождении через него пары электронов создается градиент концентрации протонов через мембрану тилакоидов, направленный внутрь мембраны, - рН внутри тила-коида может достигать значений 4 и ниже. [13]
Распад тирозина начинается с трансаминирования, приводящего к образованию и-оксифенилпировиноградной кислоты. Последняя при действии фермента оксидазы и-оксифенилпировиноградной кислоты ( медьсодержащий белок) катализирует превращение своего субстрата в гомогентизиновую кислоту, причем реакция эта включает в себя декарбоксилирование, окисление, гидроксилирование ароматического ядра и миграцию боковой цепи. Оксидаза гомогентизиновой кислоты - фермент, нуждающийся в ионах двухвалентного железа, катализирует образование из гомогентизиновой кислоты малеилаце-тоуксусной кислоты. Последняя превращается в соответствующее транссоединение - фумарилацетоуксусную кислоту, расщепление которой приводит к образованию, во-первых, фумаровой кислоты, которая подвергается дальнейшим превращениям при участии ферментов цикла ТКК, и, во-вторых, ацето-уксусной кислоты, которая может либо войти также в цикл ТКК ( в форме аце-тил - КоА), либо превратиться в жирные кислоты. [14]
Электрон от акцептора фотосистемы II проходит через цепь переносчиков и поступает в реакционный центр фотосистемы I, на фотоокисленную форму хлорофилла а - пигмент П700 ( ii0 500 мВ), заполняя электронную вакансию аналогично тому, как это происходит при фотосинтезе зеленых серобактерий. Перенос электронов от акцептора электронов фотосистемы II до реакционного центра фотосистемы I - темновой процесс, состоящий из серии этапов, в которых участвуют переносчики с понижающимися восстановительными потенциалами, такие как ци-тохромы разного типа, пластоцианин ( медьсодержащий белок), пластохинон. Электронный транспорт на этом участке на определенных этапах сопровождается ориентированным поперек мембраны переносом протонов и, следовательно, генерированием Дрн разрядка которого с помощью протонной АТФ-синтазы приводит к синтезу АТФ. [15]
Страницы: 1 2