Белок-активатор
Cтраница 1
 |
Образование петли ДНК при кооперативном взаимодействии рспрессора с операторными участками, разделенными целым числом витков спирали ДНК. [1] |
Белок-активатор катаболитных оперонов ( БАК) в комплексе с циклическим сАМР активирует транскрипцию большого числа оперонов, отвечающих за расщепление различных соединений, преимущественно Сахаров, используемых бактериальной клеткой в качестве источникон энергии и углерода. Концентрация с AMP в клетках повышается при росте на плохо усваиваемых источниках, например ацетате или глицерине, и снижается при росте ни легко усваиваемых, например глюкозе. Поэтому система регуляции с помощью БАК-сАМР позволяет клетке включать опероны катаболизма лишь по мере истощения более легко усваиваемых пищевых веществ. БАК состоит из двух идентичных субъединиц. Каждая субъеди-ница образует два домена, но в отличие от репрессора фага Я за связывание с ДНК отвечает С-концевой домен. N-концевой домен БАК связывается с сАМР и обеспечивает межсубъединичные контакты в димере. [2]

Белок-активатор катаболитных оперонов ( БАК) в комплексе с циклическим сАМР активирует транскрипцию большого числа оперонов, отвечающих за расщепление различных соединений, преимущественно Сахаров, используемых бактериальной клеткой в качестве источников энергии и углерода. Концентрация с AMP в клетках повышается при росте на плохо усваиваемых источниках, например ацетате или глицерине, и снижается при росте на легко усваиваемых, например глюкозе. Поэтому система регуляции с ломощью БАК-сАМР позволяет клетке включать опероны катаболизма лишь по мере истощения более легко усваиваемых пищевых веществ. БАК состоит нз двух идентичных субъединиц. Каждая субъеди-яица образует два домена, но в отличие от репрессора фага Я за связывание с ДНК отвечает С-концевой домен. N-концевой домен БАК связывается с сАМР и обеспечивает межсубъединичные контакты в димере. [4]

Белок-активатор катаболитных оперонов ( БАК) в комплексе с циклическим сАМР активирует транскрипцию большого числа оперонов, отвечающих за расщепление различных соединений, преимущественно Сахаров, используемых бактериальной клеткой в качестве источников энергии и углерода. Концентрация с AMP в клетках повышается при росте на плохо усваиваемых источниках, например ацетате или глицерине, и снижается при росте на легко усваиваемых, например глюкозе. Поэтому система регуляции с помощью БАК-сАМР позволяет клетке включать опероны катаболизма лишь по мере истощения более легко усваиваемых пищевых веществ. БАК состоит из двух идентичных субъединиц. Каждая субъединица образует два домена, но в отличие от репрессора фага Я за связывание с ДНК отвечает С-концевой домен. N-концевой домен БАК связывается с сАМР и обеспечивает межсубъединичные контакты в димере. [6]
Простейший механизм репрессии заключается в стерическом блокировании репрессором присоединения РНК-полимеразы к промотору. Такой механизм имеет место в тех промоторах, в которых участок связывания репрессора перекрывается с участком связывания РНК-полимеразы. Простейший механизм активации заключается в том, что белок-активатор присоединяется к промотору рядом с РНК-полимеразой и за счет непосредственного контакта с ней облегчает образование открытого промоторного комплекса. Дискуссионными являются механизмы действия тех белков-регуляторов, которые присоединяются к ДНК на значительном расстоянии от РНК поли-меразы. Ниже рассмотрено несколько наиболее хорошо изученных примеров, иллюстрирующих различные принципы регуляции промоторов. [7]
Простейший механизм репрессии заключается в стерическом блокировании репрессором присоединения РНК-пол имеразы к промотору. Такой механизм имеет место в тех промоторах, в которых участок связывания репрессора перекрывается с участком связывания РНК-пол имеразы. Простейший механизхМ активации заключается в том, что белок-активатор присоединяется к промотору рядом с РНК-полимеразой и за счет непосредственного контакта с ней облегчает образование открытого промоторного комплекса. Дискуссионными являются механизмы действия тех белков-регуляторов, которые присоединяются к ДНК на значительном расстоянии от РНКгПОЛи-меразы. Ниже рассмотрено несколько наиболее хорошо изученных примеров, иллюстрирующих различные принципы регуляции промоторов. [8]
Простейший механизм репрессии заключается в стерическом блокировании репрессором присоединения РНК-полимеразы к промотору. Такой механизм имеет место в тех промоторах, в которых участок связывания репрессора перекрывается с участком связывания РНК-полимеразы. Простейший механизм активации заключается в том, что белок-активатор присоединяется к промотору рядом с РНК-полимеразой и за счет непосредственного контакта с ней облегчает образование открытого промоторного комплекса. Дискуссионными являются механизмы действия тех белков-регуляторов, которые присоединяются к ДНК на значительном расстоянии от РНКгполи-меразы. Ниже рассмотрено несколько наиболее хорошо изученных примеров, иллюстрирующих различные принципы регуляции промоторов. [9]
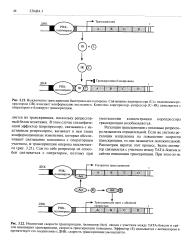
Регуляция транскрипции с помощью репрессо-ра называется отрицательной. Если же система регуляции направлена на повышение скорости транскрипции, то она называется положительной. Рассмотрим вкратце этот процесс. Белок-активатор связывается с участком между ТАТА-боксом и сайтом инициации транскрипции. [11]
Синтез мРНК и соответственно синтез белка должны строго регулироваться, поскольку у клетки недостаточно ресурсов для одновременной транскрипции и трансляции всех структурных генов. И про -, и эукариоты постоянно синтезируют только те мРНК, которые необходимы для выполнения основных клеточных функций. У прокариот транскрипция инициируется связыванием РНК-по-лимеразы с последовательностями ТАТА и TTGAC промоторной области структурного гена или оперона. Включение или выключение некоторых оперонов осуществляется при участии эффектора, который изменяет конформа-цию белка-репрессора и препятствует блокированию транскипции. При уменьшении концентрации эффектора в клетке репрессор связывается с участком ДНК, примыкающим к сайту инициации транскрипции, и препятствует перемещению РНК-полимеразы вдоль молекулы ДНК, блокируя таким образом транскрипцию. В других оперонах с участком ДНК, соседствующим с сайтом инициации траскрипции, связывается белок-активатор, который увеличивает скорость транскрипции. Связывание эффектора с активатором может снижать скорость транскрипции. [12]
Страницы: 1