Ассимиляционная нить
Cтраница 1
 |
Хордария плетевидная ( Chordaria flagelli-f ormis. [1] |
Ассимиляционные нити располагаются пучками. Центральная часть ветвей сложена пучком нитей, слабо соединенных между собой. [2]
 |
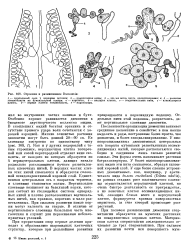
Строение Lemanea. [3] |
От этих нитей могут развиваться дополнительные пучки ветвей ограниченного роста, которые вместе с первыми служат ассимиляционными нитями. [4]
У коринофлеевых слоевище шаровидное, внутренняя часть его образована дихотомиче-ски-ветвящимися нитями, построенными из крупных бесцветных клеток. Снаружи располагаются ассимиляционные нити, а периферические нити, в отличие от элахистовых, отсутствуют. Она может достигать в диаметре 5 см. В крупных слоевищах внутренние нити разрушаются, и слоевища превращаются в полые шары. На одном слоевище могут одновременно развиваться одногнездные и однорядные многогнездные спорангии. Растет леатезия на камнях и на водорослях в литорали и в верхней сублиторали морей умеренного пояса. [5]
Середина воздушных камер у маршанции, коноцефалума и маршанциевых других родов заполнена короткими, часто ветвистыми тяжами, клетки которых содержат хлоропласты. У некоторых представителей в воздушных камерах отсутствуют ассимиляционные нити, и их функцию выполняют богатые хлороплас-тами клетки стенок камер. [6]
 |
Строение и размножение Dumontia. [7] |
По сложности организации дюмонтия занимает срединное положение в семействе: в нем можно выделить и роды, построенные более просто, чем дюмонтия, и более специализированные. У низкоорганизованных дюмонтиевых центральная ось покрыта мутовками разветвленных ассимиляционных нитей, которые располагаются свободно и соединены лишь сильно развитой слизью. Эти формы очень напоминают растения батрахоспермума. У более высокоорганизованных видов эти нити смыкаются, образуя сплошной коровой слой. У еще более сложно устроенных дюмонтиевых, как, например, у арктического вида дилсеи съедобной ( Dilsea. [8]
Представители этого семейства, близкого к гельминтокладиевым, построены также по многоосевому принципу. Главные их отличия заключаются в том, что поверхностные клетки ассимиляционных нитей слагаются в сплошной коровой слой, а нити гонимобласта окружены оберткой из тонких перепутанных нитей, в результате чего формируется мешковидный кон-цептакул с отверстием на вершине для выхода карпоспор. Кроме того, семейство характеризуется наличием обызвествленных форм. Среди них широко известна галаксаура ( Galaxau-га) - одна из самых обычных водорослей в тропических морях Мирового океана. В большей или меньшей степени обызвествленное и виль-чато-разветвленное слоевище галаксауры имеет весьма характерный облик. Любопытная особенность видов этого рода ( а их описано около 70) заключается в том, что гаметофит и спорофит, будучи оба макроскопическими растениями, заметно различаются по анатомическому строению. Это служило поводом для описания их как отдельных видов, и к настоящему времени систематика этого рода изрядно запутана. [9]
Слоевище элахистовых имеет вид маленького пушистого или плотного шарика, несущего густой пучок нитей. Внутри слоевище состоит из вильчато-ветвящихся коротких нитей, которые у поверхности переходят в неразветвленные короткие ассимиляционные и длинные периферические нити; последние при сильном развитии придают слоевищу вид пучка нитей. У некоторых представителей ассимиляционные нити отсутствуют. Органы размножения развиваются на основных нитях рядом с ассимиляционными или на периферических нитях. Большинство элахистовых растет в качестве эпифитов на крупных водорослях, некоторые имеют снизу бесцветные нити, внедряющиеся в ткань хозяина. Распространены элахистовые в холодных и умеренных морях. [10]
Слоевище у хордариевых крупное, высотой до 70 см, кустистое, с цилиндрическими ветвями. В центральной части побегов располагается пучок продольных нитей или одна нить. Поверхность слоевища образована ассимиляционными нитями. В отдельных районах морей СССР широко распространены виды пяти родов. [11]
Род аналипус ( Analipus) удобно рассмотреть на примере наиболее известного вида аналипу-са японского ( A. В СССР это довольно обычное растение в некоторых районах Японского моря, у Сахалина, Курильских островов и восточного побережья Камчатки. Аналипус японский имеет вертикальное слоевище высотой до 30 см с центральным побегом, от которого отходят короткие неветвящиеся веточки. Коровой слой образован ассимиляционными нитями. Многогнездные вместилища развиваются путем преобразования клеток ассимиляционных нитей, кроме верхушечной клетки. Вертикальное слоевище прикрепляется к грунту подошвой. Иногда оно отходит от стелющихся разветвленных многорядных побегов с плотным коровым слоем. В местах с сильным прибоем вертикальные побеги образуют небольшое число коротких боковых побегов. Могут вырастать вертикальные побеги совершенно без боковых ветвей. В еще более прибойных местах развиваются только стелющиеся побеги или слоевища в виде корок, как у ральфсии. [12]
Бесполое размножение у каулерповых отсутствует, по-видимому, оно было утрачено у современных форм в процессе эволюции. Половое размножение - анизогамия - характеризуется рядом особенностей, отличающих каулерповых от других семейств порядка. Гаметы образуются непосредственно в ассимиляционных нитях в любой их части. На отдельных участках цитоплазма сгущается, приобретает темно-зеленую окраску, затем сетчатое строение и наконец делится с образованием одноядерных гамет. Какие-либо перегородки, отделяющие место образования гамет, отсутствуют. Такое размножение называют г о л о к а р п и е и. Для выхода гамет на поверхность слоевища образуются довольно длинные выросты - папиллы; гаметы освобождаются в результате разрыва оболочки на вершинах папилл. [13]
 |
Строение и размножение Nemalion. [14] |
После оплодотворения кар-погон делится поперечной перегородкой. Из двух образовавшихся клеток только верхняя участвует в образовании нитей гонимобласта. Нижняя клетка сливается с остальными клетками карпогонной ветви, включая несущую, в одну крупную, богатую содержимым клетку слияния, служащую источником питания для развивающегося гонимобласта. У других водорослей семейства нити гонимобласта стелются между ассимиляционными нитями, образуя негустые пучки веточек с карпоспорами на конце. Карпоспоры у немалиона конечные. [15]
Страницы: 1 2