Нуклеотидом
Cтраница 3
Гены, кодирующие тРНК, значительно меньше, чем гены, кодирующие полипептиды, поскольку один нуклеотид тРНК кодируется одним нуклеотидом ДНК. [31]
Конформационный полиморфизм одноцепочечной ДНК, SSCP ( Single-strand conformational polymorphism) Различие в конформации одноцепочечных ДНК, отличающихся одна от другой всего одним нуклеотидом. Анализируемые ДНК подвергают денатурации. Денатурированные цепи принимают разную конформа-цию и при гель-электрофорезе мигрируют с разной скоростью. [32]

Полисахариды клеточной стенки обычно образуются в аппарате Гольджи и выводятся путем экзоцитоза, но из этого правила есть одно важное исключение: целлюлоза у большинства растений синтезируется на внешней поверхности клеток с помощью мембраносвязанного ферментного комплекса, субстратом для которого служит соединение сахара с нуклеотидом, вероятно UDP-глюкоза. Новообразованные целлюлозные цепи спонтанно объединяются в микрофибриллы, которые затем включаются в сложную структуру клеточной стенки. [34]
Как и ожидалось, 18О отсутствовал в изомере А, использованном в ферментативной реакции, но обнаружен в эпимере В. Следовательно, потеря 18О нуклеотидом в реакции, с инверсией, требует, чтобы введение 18О происхо-с инверсией. Эти результаты исключают псевдовращение как стадию расщепления связи и помогают определить положение, которое Н2О может занять в моделях, полученных при кристаллографических исследованиях РНазы А. [35]

Чтобы обеспечить образование непрерывной цепи ДНК из многих таких фрагментов, в действие вступает особая система репарации ДНК, удаляющая РНК-затравку и заменяющая ее на ДНК. У бактерий РНК-затравка удаляется нуклеотид за нуклеотидом благодаря 5 - З - экзонуклеазной активности ДНК-полимеразы. Завершает весь процесс фермент ДНК-лигаза, катализирующий образование фосфодиэфирной связи между группой З - ОН нового фрагмента ДНК и 5 -фосфатной группой предыдущего фрагмента. [37]
Первым выделенным мононуклеотидом была инозиновая кислота ( IMP, 9), которая была получена из гидролизата мяса Ли-бихом в 1847 г. примерно за 20 лет до выделения нуклеиновых кислот из гнойных клеток Мишером. Инозиновая кислота не является широкораспространенным в природе нуклеотидом; она образовалась в процессе выделения по Либиху за счет дезаминирования AMP, который сам по себе был выделен из мышц лишь в 1927 г. Были выделены все обычные нуклеотиды аденина, цитозина, гуанина, тимина и урацила, так же как и многие минорные нуклеотиды, например, образующиеся из псевдоуридина, дигидроуридина и метилированных производных аденозина и гуанозина. [38]
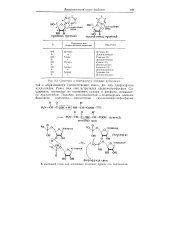 |
Структура и номенклатура основных нуклеозидов. [39] |
Со -, состоящее из основания, сахара и фосфата, называется нуклеотидом. [40]
Реакции ферментативного фосфорилирования с участием аде-нозинтрифосфата ( АТР) протекают в присутствии ионов магния. Хотя в этих реакциях ион металла может выступать как хела-тирующий агент между нуклеотидом и ферментом, ряд факторов указывает на то, что функции магния могут быть связаны с рассматриваемым здесь суперкислотным катализом. Очевидно, что при неферментативном гидролизе АТР, а также более простых полифосфатов и трифосфатов ускорение гидролиза различными ионами двухвалентных металлов - кальция, магния, марганца ( П), меди ( П) и кадмия - связано с суперкислотным катализом. Двухвалентные металлы также катализируют неферментативное трансфосфори-лирование АТР и у-фенилпропилтрифосфата с неорганическим фосфатом и карбоксилат-ионами. При некатализируемом гидролизе уФенилпРопилДиФосФата монопротонированные частицы реагируют примерно в 2000 раз быстрее, чем полностью ионизованные частицы. Образование метафосфат-иона вновь ускоряется хелатированным ионом металла, проявляющего суперкислотные свойства. Идентичность констант Скорости катализируемых ионами металлов реакций у-фенкл-пропилтрифосфата и АТР с неорганическим фосфатом показывает, что остаток аденина не связывается с ионом металла. Методом ЯМР было показано, что ионы магния, кальция и цинка преимущественно взаимодействуют с Р - и 7 - Ф сФатами АТР, ионы двухвалентной меди - с а - и р-фосфатами, а ион двухвалентного марганца связывается со всеми тремя. [41]
Поскольку карбобензоксиаминокислоты и нуклеотиды сильно различаются по своему нуклеофильному характеру, смешанные ангидриды получаются с довольно высокими выходами, а динуклео-зидпирофосфат - лишь в незначительном количестве. При атаке аддукта карбоксил-ионом может получиться ангидрид карбобензоксиаминокислоты, который затем быстро вступил бы в реакцию с нуклеотидом ( или любым динуклеозид-5 - пирофосфатом) также с образованием смешанного ангидрида. [42]
ДНК и различна даже у различных индивидуумов одного и того же вида. Одно звено ДНК, содержащее по одной группе фосфорной кислоты, сахара и азотистого основания, называется нуклеотидом. [43]
Ген заканчивается двойным стоп-сигналом UAAUAG. Далее следует межгенная последовательность из 36 нуклеотидов, лосле которой кодоном AUG начинается очень длинный ген репликазы. Этот ген заканчивается 3395 - м нуклеотидом, после которого со стороны З - конца следует нетранслируемый участок из 174 нуклеотидов. [44]
В простейших случаях взаимодействие кодона с антикодоном происходит путем образования трех канонических пар гетероциклов. Антикодон, специфичный в фенилаланину, имеет структуру GmpApA ( символ m справа от G означает метилирование 2 -окси-группы), комплементарную фенилаланиновому кодону UpUpC. Однако в большом числе случаев соответствие между третьим нуклеотидом кодона и первым нуклео-тидом антикодона имеет более сложный характер. Например, одна и та же фени-лаланиновая тРНК опознается обоими кодонами - и полностью комплементарным UpUpC и не полностью комплементарным UpUpU. Некоторые тРНК, специфичные к аминокислотам, кодируемым тремя и более кодонами, содержат в первом положении антикодона инозин. Такие тРНК могут взаимодействовать сразу с тремя кодонами, имеющими в третьем положении уридин, цитидин или адено-зин. [45]
Страницы: 1 2 3 4