О-антиген
Cтраница 2
Штаммы дикого типа с полным О-ан-тигенным ЛПС формируют гладкие колонии, тогда как мутанты или диссоцианты, лишенные О-антигенов, образуют шероховатые колонии. В некоторых шероховатых мутантах О-антиген у эн-теробактерий заменен на другой полисахарид, общий антиген энтеробактерий ( ОАЭ), широко распространенный в этой группе микроорганизмов. ОАЭ является гетерополисахаридом, состоящим из М О-диацетилглюкозамина, N-ацетиламиноманнуроновой кислоты и N-ацетилфукозамина, он консервативен среди энтеробактерий. ОАЭ может существовать в двух формах - первая прикреплена к наружному слою внешней мембраны редуцирующим концом через остаток фосфатидила и присутствует постоянно, вторая замещает О-цепи ЛПС у мутантных штаммов. [16]
Эндотоксин, следовательно, является структурным компонентом О-соматического антигена. Полисахаридная часть ЛПС определяет серологическую специфичность О-антигена, что широко используется для таксономических целей. [17]
В состав клеточной стенки грамотрицательных бактерий входит сложный липополисахарид, с которым в значительной степени связаны биологические особенности этого класса бактерий. Липополисахарид иммунологически активен ( он известен под названием О-антигена); комплекс его с белком представляет из себя эндотоксин грамотрицательных бактерий, имеющий существенное значение в процессах бактериальной инфекции. [18]
Липополисахарид ( ЛПС), обладающий свойствами эндотоксина, является одним из компонентов клеточной стенки грамотрицатель-ных бактерий. Он образует с белками и липидами сложный макро-молекулярный комплекс, обозначенный как О-антиген. [19]
Для типирования по - антигену в один ряд пробирок с разведениями диагностической ОК-сыворотки вносят суспензию живой ( негретой) культуры, а для определения О-антигена в другой ряд пробирок вносят гретую ( кипяченную в течение 1 - 2ч) взвесь бактерий. Кипячение или автоклавирование разрушает поверхностно расположенный - антиген, который мешает определению О-антигена. Через сутки инкубирования при 37 С гомологичные сыворотке штаммы агглютинируются до титра или до половины титра. [20]
Другие фрагменты в липополи-сахариде могут отличаться в зависимости от вида бактерий. Им-муногенные свойства проявляют боковые полисахаридные цепи и сердцевина. Боковые полисахаридные цепи отвечают за антигенную специфичность молекулы липополисахарида и называются О-антигенами. [21]
Кауфман [30] и Уайт [31] с большой тщательностью изучили антигенную структуру Salmonella. Разработанная этими авторами классификация Salmonella, основанная на определении соматических и жгутиковых антигенов, стала общепринятой. По этой классификации все виды Salmonella разделены на ряд групп, принадлежность к той или иной группе определяется сходством О-антигенов. Не следует, однако, забывать, что вся классификация построена на чисто серологических данных. [22]
 |
Структура липополисахарида Salmonella typhimurium. [23] |
ЛПС образуют на поверхности клетки отрицательный заряд и часто являются токсичными. Структура Kdo-липида А высоко консервативна среди грамотрицательных микроорганизмов. Центральная оли-госахаридная область также до некоторой степени консервативна, а у представителей энтеробактерий - похожа, хотя и не идентична. О-антигены содержат длинные полисахариды, состоящие из повторяющихся олигосахаридных единиц. Характерные структуры могут содержать дезокси -, дидезокси - и аминосахара. [24]
ЛИПОПОЛИСАХАРИДЫ, соединения, макромолекулы к-рых состоят из трех частей, называемых соотв. Липид А построен из фосфорилиров. D-глюкозаминил - ( 1 - 6) - О-глюкозамина, в к-рых гидроксилы и аминогруппы ацилированы высшими жирными к-тамн. Кор - олиго-сахарид, и состав к-рого наряду с обычными гексозами и гек-созаминами входят гептозы, 2-кето - З - дезоксиоктоновая к-та и fl - аминоэтилфосфат ( этаноламинфосфат); одинаковый по структуре кор могут иметь сравнительно большие группы бактерий. О-антигены), являются сильными эндотоксинами. ЛИПОПРОТЕИДЫ ( липонротеины), комплексы, содержащие липиды и белки, связь между к-рыыи осуществляется посредством гидрофобных и электростатич. К последним относятся также и протеолипиды, к-рые раств. ЛОНП ( 64 % ацилглицеринов и эфиров холестерина, 20 % фосфолипидов и 8 % белка; плотн. ЛОНП); хиломикроны остаются на старте. [25]
ЛИПОПОЛИСАХАРИДЫ, соединения, макромолекулы к-рых состоят из трех частей, называемых соотв. Липид А построен из фосфорилиров. D-глюкозаминил - ( 1 - 6) - О-глюкозамива, в к-рых гидроксилы и аминогруппы ацилированы высшими жирными к-тами. Кор - олиго-сахарид, в состав к-рого наряду с обычными гексозами и гек-созаминами входят гептозы, 2-кето - З - дезоксиоктоновая к-та и р-аминоэтилфосфат ( этаноламинфосфат); одинаковый по структуре кор могут иметь сравнительно большие группы бактерий. О-антигены), являются сильными эндотоксинами. ЛИПОПРОТЕИДЫ ( липопротеины), комплексы, содержащие липпды и белки, связь между к-рыми осуществляется посредством гидрофобных и электростатич. К последним относятся также и протеолипиды, к-рые раств. ЛОНП ( 64 % ацилглицеринов и эфиров холестерина, 20 % фосфолипидов и 8 % белка, плотн. [26]
Она может быть полипептидной, рибитфосфатной или состоять из смешанных полимеров. Определяющее значение для биологической функции имеют концевые олигосахаридные цепи, в состав которых обычно не входят уроновые кислоты или сульфоэфиры. Носителем отрицательных зарядов являются лишь остатки сиаловых кислот. Примерами таких соединений могут служить групповые вещества крови, муцин подчелюстной железы, О-антиген грамотрицательных бактерий, полисахарид капсулы пневмококков типа XIV, декстраны. [27]
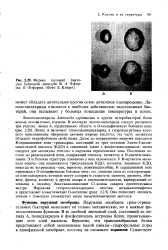 |
Формы колоний бактерии Salmonella paratypki В. Л-8-фор-ма. B-R - форма. ( Фото Е. Kroger. [28] |
Липополисахариды Salmonella typhimurium и других энтеробактерий были весьма основательно изучены. Липид А включает глюкозаминдисахарид, к гидроксигруппам которого присоединены эфирными связями жирные кислоты; эта часть молекулы обладает гидрофобными свойствами. Далее по направлению кнаружи находится R-сердцевинная зона-трисахарид, состоящий из трех остатков 2-кето - З - дезок-сиоктоновой кислоты ( КДО) и связанный также с фосфоэтаноламином; затем следуют две молекулы гептозы и наружная сердцевинная зона. Последняя состоит из разветвленной цепи, содержащей глюкозу, галактозу и N-ацетилглюко-замин. Эта базовая структура одинакова у всех сальмонелл. У R-мутантов молекула короче-она оканчивается трисахаридом КДО; мутанты, лишенные КДО, никогда не встречались - они, вероятно, нежизнеспособны. Восстанавливающие С1 - концы сахароз ориентированы внутрь. Наружная гетерополисахаридная цепочка штаммоспецифична и представляет ( соматические) О-антигены ( разд. Она позволяет опознать штамм с помощью упомянутых выше иммунохимических методов. [29]
Страницы: 1 2