Энергетический обмен - клетка
Cтраница 1
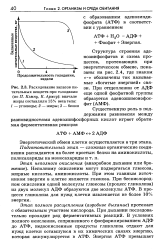 |
Расходование запасов питательных веществ при голодании ( по 77. Кэмпу, К. Армсу. вначале жиры составляли 15 % веса тела. 1 - углеводы. 2 - жиры. 3 - белки. [1] |
Энергетический обмен клетки осуществляется в три этапа. [2]
При изучении регуляции энергетического обмена клетки отправным пунктом, на котором обычно строится исследование, является открытый Пастером феномен подавления менее эффективного в энергетическом отношении брожения более эффективным дыханием. Выяснению механизма этого регуляторного феномена посвящены многочисленные глубокие исследования и покоящиеся на этих исследованиях плодотворные гипотезы. Весьма существенно, однако, что объектом такого рода исследований обычно служат переживающие in vitro интактные клетки - взвеси свободных клеток или срезы тканей. Эти интактные клетки в момент исследования в функциональном отношении находятся в состоянии относительного покоя, и очевидно вследствие этого пастеровский эффект () выражен у них в полной мере. Если же проследить метаболизм таких же клеток или клеток других животных тканей в условиях активно функционирующего целого организма, то оказывается, что их энергетический обмен характеризуется не пастеровским торможением гликолиза, а как раз обратным состоянием - сосуществованием дыхания иаэробного гликолиза. Многие авторы проходят мимо этого факта, хотя хорошо известно, что все ткани животного организма при напряженной работе in situ обнаруживают наряду с повышенным поглощением кислорода высокий аэробный гликолиз, иногда в 1000 раз превосходящий по скорости тот уровень гликолиза, который отмечается в покоящейся ткани. [3]
Главная роль в энергетическом обмене клеток животных принадлежит дыхательному обмену или клеточному дыханию. Клеточное дыхание представляет собой процесс, в котором высокомолекулярные органические высокоэнергетические соединения, окисляясь распадаются на низкомолекулярные или неорганические соединения, бедные энергией. При окислении с участием кислорода дыхание называют аэробным, а без его участия - анаэробным. [4]
Активность многих ферментов, участвующих в энергетическом обмене клетки, по крайней мере частично регулируется энергетическим статусом клетки, причем главными метаболическими сигналами служат аденилаты. В мышце устрицы реакции, катализируемые обоими интересующими нас ферментами, ведут к образованию высокоэнергетических фосфатов ( АТФ в случае пируваткиназы и ИТФ или ГТФ - в случае фосфоенолпируваткарбоксикиназы), и обе они подвержены ингибированию своим продуктом. В этом отношении оба фермента ведут себя в соответствии с концепцией энергетического заряда Эткин-сона. Такой механизм, несомненно, способствует физиологическому равновесию в точке разветвления пути фосфоенолпирувата, но сам по себе он не обладает достаточной специфичностью, чтобы им можно было объяснить переход от аэробного обмена к анаэробному. Необходимая специфичность может быть привнесена ионами Н и L-аланином. [5]
ФК ( см. рис. 4) восстанавливает энергетический обмен клеток печени и обеспечивает, несмотря на дефицит инсулина и значительную степень восстановления, включение метки в белки. В то же время эти результаты показывают, что нарушение включения меченой аминокислоты в белки печени крыс с аллоксановым диабетом в основном обусловлено не выпадением прямого действия инсулина на синтез белка, а зависит от снижения энергетического обеспечения этого синтеза вследствие блока глюкокиназной реакции. [7]
Креатинкиназа - фермент, принимающий участие в энергетическом обмене клеток мышечной, нервной и других тканей. Повышение активности креатинкиназы наблюдается при повреждении миокарда ( инфаркт миокарда, миокардит), мышечной ткани ( травма, мышечные дистрофии, дерматомиозит, полимиозит и др.), при обширных оперативных вмешательствах, тяжелой физической нагрузке, внутримышечных инъекциях лекарственных средств ( особенно наркотических и обезболивающих), гипотиреозе, острой алкогольной интоксикации. [8]
Таким образом, АТФ играет центральную роль в энергетическом обмене клетки. [9]
Соединения фосфора с адениловой кислотой занимают ведущее место в энергетическом обмене клетки. Они содержат фосфатные связи, в которых аккумулируется энергия, освобождающаяся в. В то же время поглощение и усвоение фосфора определяется наличием в растениях других питательных элементов, и в первую очередь азота. [10]
Совокупность механизмов получения энергии за счет внешних энергетических ресурсов и путей дальнейшего расходования ее клеткой при осуществлении многообразных жизненных функций составляет энергетический обмен клетки. [11]
Тироксин, трийодтиронин, трийодтироуксусная кислота и некоторые другие йодированные соединения, образуемые щитовидной железой, резко усиливают окислительные процессы. В наибольшей мере активизируются окислительные процессы в митохондриях, что ведет к усилению энергетического обмена клетки. [12]
В настоящее время изучена тонкая структура ( ультраструктура) этих элементов. В наружных члениках палочек и колбочек сосредоточены диски, осуществляющие фотохимические процессы, на что указывает повышенная концентрация родопсина в дисках палочек и йодопсина в дисках колбочек. К наружным сегментам палочек и колбочек прилежит скопление митохондрий, которым приписывается участие в энергетическом обмене клетки. Палочконесущие зрительные клетки являются аппаратом сумеречного зрения, колбочконесущие клетки - аппаратом центрального и цветового зрения. [13]
Такой взгляд на фенолъные соединения, который, разумеется, не способствовал более глубокому их изучению, был решительно отвергнут классическими работами академиков В, И. Созданные этими учеными теории Дыхательных хромогенов ( В. И. Палладий) и Медленного биологического окисления ( А. Н. Бах), получившие мировбе признание, отводили полйфеноМм исключительно а & тйвную роль в энергетическом обмене клетки, что заставило по-новому взглянуть на эту группу соединений. Открываются все новые аспекты их участия в физиологических процессах и их практического применения. [14]
На проницаемость мембраны могут влиять различные факторы. Так, инсулин повышает проницаемость плазматической мембраны мышечных клеток для глюкозы, стимулируя транспорт глюкозы из крови и межклеточных пространств внутрь клеток скелетной и сердечной мышцы и жировой ткани. При интенсивном течении процессов окислительного фосфо-рилирования, приводящих к накоплению больших количеств АТФ, внутри митохондрий происходит взаимодействие АТФ с актомиозинподобным белком мембран, сопровождающееся конформационными изменениями белка. С уменьшением концентрации АТФ внутри митохондрий проницаемость мембран увеличивается. По-видимому, митохондриальная мембрана участвует в регуляции энергетического обмена клетки. [15]
Страницы: 1