Образование - зигота
Cтраница 1
Образование зигот в процессе конъюгации может происходить у мезотениевых тремя путями. Клеточные оболочки в месте соприкосновения растворяются или разрываются, и клеточное содержимое из обеих клеток выходит через конъюгационный канал. [1]
В первые минуты образования зиготы в ней появляются многочисленные сокращающиеся вакуоли. Маленькие вакуоли сливаются в более крупные. Каждая вакуоля пульсирует в течение 4 - 7 мин, затем сжимается, и ее содержимое выходит наружу. Действие пульсирующих вакуолей длится от 4 до 32 мин, до тех пор пока прозрачные места зиготы перестают быть видимыми. За этот период ( от 1 до 3 5 ч) зигота значительно сокращается. Внутри ее хлоро-пласты и пиреноиды располагаются в периферическом слое. Затем начинают появляться первые признаки орнаментации зиготы - конусовидные бугорки, разбросанные по ее поверхности. Постепенно они вытягиваются и приобретают вид шипов. За время разрастания шипов, которое продолжается 2 - 3 ч, зигота опять увеличивается в размерах и становится такой же, как при слиянии гамет. [2]
Конститутивность проявлялась при образовании зиготы, но только по отношению к тем цистронам, которые находятся в положении cis относительно точки ос. [3]
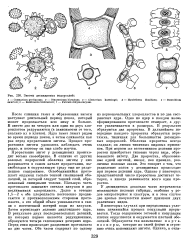 |
Зиготы десмидиевых водорослей. [4] |
После слияния гамет и образования зиготы наступает длительный период покоя, который может продолжаться всю зиму и больше. Ядра гамет лежат рядом во время периода покоя, а затем сливаются как раз перед прорастанием зиготы. Процесс прорастания зиготы удавалось наблюдать очень редко, и поэтому он еще слабо изучен. [5]
Необходимо отметить, что образование зигот у конъюгат - явление довольно редкое, а у многих видов десмидиевых и мезотепиевых водорослей зиготы вообще еще не найдены. В большинстве случаев одноклеточные копъюгаты размножаются обычным вегетативным делением в поперечной плоскости, а нитчатые - распадением нитей на отдельные клетки. [6]
У десмидиевых может происходить также образование двойных зигот. У видов клостериума способ образования двойных зигот иной: в каждой клетке сначала развиваются две особые гаметы, которые, сливаясь, дают двойные зиготы. Очень редко в образовании зигот десмидиевых принимают участие три или четыре особи. [7]
При половом размножении происходит слияние двух гамет с образованием зиготы, из которой развивается новый организм. Половое размножение ведет к генетической изменчивости. Генетическая изменчивость выгодна виду, так как она поставляет сырье для естественного отбора, а следовательно, и для эволюции. Потомки, оказавшиеся наиболее приспособленными к данной среде, будут обладать преимуществом в конкуренции с другими представителями того же вида и иметь больше шансов выжить и передать свои гены следующему поколению. Повышение изменчивости может быть достигнуто путем смешения генов двух разных особей - процесса, называемого генетической рекомбинацией, и составляющего важную особенность полового размножения. В примитивной форме генетическая рекомбинация встречается уже у некоторых бактерий ( разд. [8]
Половое размножение представляет собой или слияние двух одинаковых соседних клеток гифы и образование зиготы в одной из них ( как у базидиоболуса из энтомофторовых грибов), или слияние равных клеток двух соседних гиф и образование зиготы в перемычке между ними или в выросте из нее. Как происходит прорастание зиготы, неизвестно. [9]
Зерно ( семя) как и живая биологическая система дышит с момента образования зиготы в течение всего периода формирования и последующего своего существования. Дыхание - процесс экзотермический, и следовательно, зерновка в результате физиологических функций генерирует тепло со времени появления эмбриона. [10]
Половым размножением называют процесс получения потомства в результате слияния двух гамет с образованием диплоидной зиготы, из которой развивается новый зрелый организм. Процесс слияния гамет называют оплодотворением. При оплодотворении сливаются ядра гамет, что приводит к объединению двух наборов хромосом, по одному от каждой родительской особи. Клетку, содержащую два набора хромосом, называют диплоидной. [11]
Гаметогенез, высвобождение и слияние мужской и женской герминативных клеток - предварительные события, ведущие к образованию зиготы. Клетки спермы, попадающие во влагалище, должны продвинуться в шейку матки и, двигаясь поступательно в полости матки, достичь маточных труб, чтобы встретиться с яйцеклеткой. Процесс оплодотворения схематически заключается в проникновении сперматозоида в яйцеклетку и слияние их ДНК. После оплодотворения начинается деление клеток, которое продолжается в течение последующих трех-четырех дней, формируя морулу. Клетки морулы продолжают делиться, продвигаясь по маточным трубам, и к моменту, когда развивающийся эмбрион достигает матки, образуют полый шарик, называемый бла-стоцистой. [12]
У многих низших растений, включая водоросли, а также у простейших редукция числа хромосом происходит сразу после образования зиготы, так что организм оказывается гаплоидным. У растений с етеро-фазной ( антитетической) сменой поколений ( мхов, папоротников) гаплоидные поколения чередуются с диплоидными. [13]
Это возникновение доминантных летальных мутаций в половых клетках насекомого обычно не мешает созреванию обработанной клетки в гамету или участию гаметы в образовании зиготы, но препятствует развитию зиготы до зрелого состояния. [14]
ОПЛОДОТВОРЕНИЕ, сивгамия, слияние мужской половой клетки ( сперматозоид, спермий) с женской ( яйцо, яйцеклетка), приводящее к образованию зиготы, к-рая дает начало новому организму. [15]
Страницы: 1 2