Образование - ацетилкофермент
Cтраница 1
 |
Схема взаимопревращений пентозофосфатов. [1] |
Образование ацетилкофермента А имеет сложный характер и начинается с присоединения аниона тиамина к карбонильной группе кислоты. При участии декарбоксилазы пировиноградной кислоты из продукта присоединения тиамина образуется активный ацетальдегид, который реагирует с ли-поевой кислотой, образуя ацетиллипоевую кислоту. Ацетильная группа присоединяется к КоА - SH и образуются ацетил-кофермент А и дигидролипоевая кислота. Последняя вновь окисляется НАД или ФАД до липоевой кислоты. [2]
Коль скоро произошло образование ацетилкофермента А, все последующие реакции, ведущие к синтезу жирных кислот, по-видимому, будут сходны с реакциями, происходящими в других биосинтетических системах. В хлоропластах в этих реакциях потребляются АТФ и восстановленный НАДФ, образовавшиеся в фотохимических реакциях фотосинтеза. [3]
Другой возможный путь образования ацетилкофермента А из продуктов углеродного цикла предполагает фосфорокластическое расщепление соединения IX ( гликольальдегидтиамин-пирофосфат), образующегося в транскетолазной реакции. [4]
Донором ацетильных групп могут служить ацетат-ионы, как уже отмечалось выше, причем образование ацетилкофермента А из этих ионов нуждается в присутствии аденозинтрифосфорной кислоты. Однако важнейшим источником ацетилкофермента А в живых организмах является пировиноградная кислота, образующаяся при гликолизе. При переносе ацетильной группы от ацетилкофермента А акцептору ацетильной группы эта энергия не теряется, так как реакция сочетается с образованием молекулы АТФ из АДФ и неорганического фосфата, как было указано в другом месте. [5]
ТДФ-зависимая пируватдегидрогеназа принимает участие в окислит, декарбоксилировании пировиноградной к-ты ( пирувата) с образованием ацетилкофермента А. При этом пируват, образующийся в результате гликолитич. Общее кол-во энергии, получаемой в результате окисления пирувата в этом цикле, почти в 4 раза превосходит энергию, освобождаемую в предшествующих р-циях гликолиза. [6]
Поэтому трудно представить, чтобы сама по себе эта реакция могла обеспечить тот исключительно быстрый переход 14С из глюкозы в глутаминовую кислоту, который происходит в тканях мозга; однако для тканей мозга характерна высокая интенсивность гликолиза, а следовательно, и высокая скорость образования ацетилкофермента А, из которого С включается в углеродные скелеты а-кетокислот цикла; г) кровоснабжение мозга ( в пересчете на единицу массы) превышает кровоснабжение мышц и печени, что обусловливает более интенсивный приток изотопа к мозгу. При интерпретации полученных данных в рассмотренной системе необходимо было принимать в расчет быстрый перенос углеродного скелета от одного соединения к другому. [7]
Ацетилкофермент А - главный продукт, завершающий вторую фазу окисления углеводов, жирных кислот, глицерина, ряда аминокислот. Механизм образования ацетилкофермента А при распаде других веществ рассмотрим при изучении их обмена, а сейчас остановимся на дальнейших путях превращений ацетилкофермента А. [8]
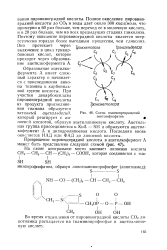 |
Схема взаимопревращений пентозофосфатов. [9] |
Полное окисление пировино-градной кислоты до СО2 и воды дает около 300 ккал / моль, что примерно в 60 раз больше, чем переход ее в молочную кислоту, и в 20 раз больше, чем на всех предыдущих стадиях гликолиза. Поэтому окисление пировиноградной кислоты является энергетически гораздо более выгодным процессом, чем гликолиз. Оно протекает через включение в цикл трикар-боновых кислот, которое проходит через образование ацетилкофермента А. [10]
Аналогично этому выяснению роли липоевой кислоты в качестве кофактора, участвующего в реакции окислительного декарбоксилирования а-кетокислот, способствовали следующие наблюдения. Во-первых, природное соединение, называемое ацетатзамещающий фактор, заменяло ацетат, необходимый для роста Lactobacillus easel, и, во-вторых, для окисления пирувата у Streptococcus faecalis необходим фактор окисления пирувата. Эти факторы содержат одно и то же соединение - липоевую кислоту, необходимую для образования ацетилкофермента А из пирувата при окислительном декарбоксилиро-вании. [11]
Широко распространены в живых организмах и играют важную роль в обмене веществ. Окислит, декарбоксилирование пи-ровиноградной к-ты под действием фермента пируватдекарбоксилазы в тканях животных и растений приводит к образованию ацетилкофермента А, вступающего в цикл трикарбоновых к-т. Эта реакция обеспечивает осн. [12]
За последнюю четверть века мы довольно много узнали о механизме восстановления углерода при фотосинтезе. И тем не менее, как это явствует из всего предыдущего обсуждения, остается еще много вопросов, которые пока не получили ответа. Так, остаются неясными многие детали реакций, ведущих от цикла восстановления углерода к различным вторичным путям биосинтеза. Требуют дальнейшего уточнения две важные проблемы - механизм образования ацетилкофермента А и гликолевой кислоты из продуктов цикла восстановления углерода. [13]
Страницы: 1