Биосинтез - фенольное соединение
Cтраница 1
Биосинтез фенольных соединений будет рассмотрен в гл. В этой главе обсуждаются в основном конечные стадии превращения этих соединений, включающие процессы гликозилирования, метилирования, этерификации и другие, снижающие их реакционную способность. Установлено, что феноль-ные соединения накапливаются в активно метаболизирующих растительных тканях в виде гликозидов или сложных эфиров. В свободном состоянии фенолы встречаются реже, хотя они могут сохраняться как таковые в специальных вместилищах масел. В растениях осуществляется регулирование процессов метаболизма фенольных соединений, в результате которых фенолы превращаются в гликозиды и другие производные. Подобные процессы в биохимической литературе обычно обобщают термином детоксикация. Весьма странным является то обстоятельство, что растения одинаково хорошо могут глико-зилировать или этерефицировать как синтетические, так и природные фенолы. Это может указывать на то, что в растениях присутствует или несколько ферментов с низкой специфичностью или значительное число высокоспецифичных ферментов. [1]
Для биосинтеза фенольных соединений могут использоваться, по-видимому, не только ацетатные, но и малонатные фрагменты. Ферментативный механизм циклизации активированных ацетатных и малонатных остатков в фенольные соединения не изучен. [2]
Гипотеза о биосинтезе фенольных соединений из-аминокислот, точнее из продуктов их распада, не получила экспериментального подтверждения в позднейших исследованиях. Хотя опыты с изотопами и показывают, что ароматические аминокислоты используются растениями для построения многих фенольных соединений ( см. ниже), такое включение пе сопровождается их расщеплением - это пример использования уже готового фенольного ядра. [3]
Степень участия фотоморфогенетической и фотосинтетической системы в биосинтезе фенольных соединений остается до сих пор слабо изученной. [4]
Помимо пути через шикимовую кислоту существует вторая возможность биосинтеза фенольных соединений - так называемый ацетатный путь. [5]
Эта глава представляет собой обзор по ферментам, участвующим в биосинтезе фенольных соединений. Подробно изучено незначительное число ферментов, и данные о их свойствах неоднозначны, хотя в природе встречается множество различных фенольных соединений. Задача до некоторой степени упрощается, если рассмотреть работы по частичной очистке ферментов ( или ферментных систем) или реакции, которые по своей стехиометрии, необходимым кофакторам и другим свойствам проходят с участием одного фермента. Почти не имеет смысла перечислять все превращения, которые отмечены в бесклеточных экстрактах. Иногда трудно провести резкое разграничение между биосинтезом и распадом. Так, например, биосинтез тирозина из фенилаланина в животных тканях является в то же время начальной стадией распада фенилаланина. [6]
Считается, что формирование отличительного вкуса и аромата не зависит от биосинтеза фенольных соединений и пигментов. Эту точку зрения подтверждает наличие темноокрашенных вин и вин с высоким содержанием таннина, обладающих слабовыраженным вкусом и ароматом. В регионах, где виноградарство только начинает развиваться, для определения оптимальной степени зрелости ягод остается использовать лишь метод проб и ошибок - в основном по определению содержания в ягодах сахара, как это и делается в большинстве регионов. Определение содержания сахара - это, скорее, простой, легко измеримый индикатор степени зрелости ягоды, а не показатель, жестко привязанный к другим, более важным компонентам состава винограда. При таком подходе важно осознавать, что он строится на предположении, что формирование желаемых компонентов вкуса и аромата в ягодах происходит параллельно накоплению в них сахара. [7]
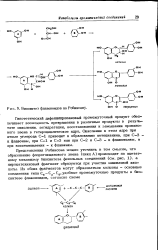 |
Биосинтез флавоноидов по Робинсону. [8] |
Представления Робинсона можно уточнить в том смысле, что образование флороглюцинового звена ( цикл А) происходит по ацетатному механизму биосинтеза фенольных соединений ( см. рис. 1), а пирокатехиновый фрагмент образуется при участии шикимовой кислоты. [9]
Фенольные соединения считаются вторичными метаболитами в растениях. Они образуются из первичных метаболитов - углеводов - по двум основным механизмам, описанным в этой главе. Ацетатный механизм приводит к образованию фенольных соединений преимущественно в низших растениях, механизм же с участием шикимовой кислоты относится главным образом к высшим растениям. Нередко оба эти пути биосинтеза фенольных соединений объединяются, как, например, при образовании флавоноидов. [10]
Биосинтез флавоноидов осуществляется в очень больших масштабах. Показано, что у высших растений на него расходуется почти 2 % всего углерода, фиксируемого при фотосинтезе. В общих чертах биосинтез флавоноидов изучен достаточно хорошо, однако многие детали, главным образом касающиеся ферментов, еще ждут своего выяснения. Удобнее всего рассматривать биосинтетический путь по стадиям: 1) образование основного Сб - С3 - Сб-скелета, включая два главных пути биосинтеза фенольных соединений - поликетидный и шикимат-ный, 2) пути, по которым флавоноиды различных классов образуются из основного Сб - С3 - Сб-предшественника, и возможные взаимопревращения между флавоноидами различных классов, 3) окончательные модификации, такие, как гидроксилирование, метилирование и гликозилирование, которые дают начало многим индивидуальным флавоноидам внутри каждого класса. [11]
 |
Центральная роль коричных кислот в биосинтезе других фенольных соединений. [12] |
Недавно Эль-Басиони и Ниш [32] опубликовали результаты изучения связанных форм коричных кислот у злаков. В отличие от свободных коричных кислот, извлекаемых ацетоном, связанные формы могут быть переведены в раствор лишь при последующем настаивании с фосфатным буфером. Оказалось, что при фотосинтезе в С1402 связанные формы метятся быстрее, чем свободные. При этом в проростках содержится несколько связанных форм коричной, и-кумаровой, кофейной и феруловой кислот. Часть связанных форм осаждается ( NH4) 2S04 и показывает молекулярный вес, приближающийся к весу белков. Авторы допускают возможность, что связанцые формы коричных кислот являются ацилированными ферментами. Они считают, что именно эти формы играют важную роль в биосинтезе фенольных соединений. [13]
Страницы: 1