Необычное основание
Cтраница 1
Необычные основания выделены из матричных РНК - 7-метилгуанозин, 1-метил - 2-амино - 6-оксопурин, 6-диметиламинопурин, обнаруженный также в природном нуклеозиде пуромицине. [1]
Имеются данные, что для s - PHK из зародышей пшеницы характерно более высокое относительное содержание необычных оснований, чем для рибосомной РНК. Соотношение оснований в s - PHK из Vigna sesquipedalis отличается от соотношения оснований в рибосомной РНК, выделенной из этой же ткани. Еще не показано, что для s - PHK из растений характерно соответствие в соотношении оснований, обнаруженное для s - PHK из других организмов. [2]
В отсутствие воды или в присутствии сильного водоотнимающего средства во фто-ро -, хлоро - или оксосистемах могут быть получены многочисленные солеобразные соединения, производные совершенно необычных оснований. [3]
Заметим, однако, что видовая специфичность биосинтеза белка тем не менее существует; проявляется она в различном узнавании аминоацилсинтетазами ( ферментами, активирующими аминокислоты) транспортных РНК; такая специфичность обусловлена, по-видимому, присутствием различных необычных оснований в s - PHK разных видов. [4]
Сообщалось, что рибосомы из проростков гороха содержат 40 % РНК, 60 % белка и 4 моль двухвалентного катиона на 1 моль основания. В рибосомной РНК необычные основания обнаружены только в очень небольших количествах. [5]
На 5 -конце молекулы тРНК всегда находится гуанин, а на З - конце - последовательность оснований ЦЦА. Последовательность нуклеоти-дов в остальной части молекулы варьирует и может содержать необычные основания, такие как инозин и псевдоурацил. Каждая аминокислота присоединяется к одной из своих специфичных тРНК при участии фермента амино-ацил - тРНК - синтетазы. [6]
Во-первых, в нее входит 5 - 9 % псевдоурацила; во-вторых, имеются метилированные пурины и пиримидины, например 6-диметиламинопурин, причем в столь малых количествах, что приходится предполагать, что эти необычные основания входят лишь в избранные типы РРНК. Присутствие некоторых специальных оснований в РРНК, специфичной к одной из аминокислот, например гистидину, связано с механизмом прикрепления и механизмом избирательности. По-видимому, специфичность в этом случае заключена в связывании каждой специальной РРНК со своим специфическим ферментом. Эта комбинация реагирует с одной определенной аминокислотой. [7]
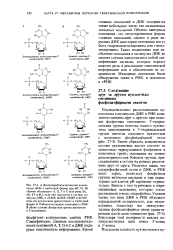
ДНК содержится также небольшое число так называемых минорных оснований. Обычно минорные основания-это метилированные формы главных оснований, однако в ряде вирусных ДНК некоторые основания могут быть гидроксиметилированы или глюко-зилированы. Такие измененные или необычные основания в молекулах ДНК во многих случаях представляют собой специфические сигналы, которые играют важную роль в реализации генетической информации или в обеспечении ее сохранности. [9]
Азагуанин - яд, тормозящий многие синтетические реакции в бактериях. В частности, он тормозит синтез белков и рибосомной РНК. Однако та специфическая РНК, которая служит матрицей для синтеза белков, продолжает образовываться в клетке и включает в себя необычное основание взамен гуанина. Если после не слишком долгого периода отравления азагуанином клетки снова переносятся на обычную среду, синтез белков и рост возобновляются, но опять, как в случае 5-фторурацила, клетка образует измененные белки вместо нормальных. [10]
Одно время считали, что роль матрицы, с которой соединяются молекулы аминоацил - РНК и на которой происходит сборка белковой молекулы, играет стабильная рибосомная РНК. Однако нуклеотидный состав ДНК очень слабо коррелирует с нуклеотидным составом рибосомной РНК. Кроме того, известно, что если какой-либо ген поврежден, бактерии теряют способность синтезировать белок. И, наконец, при включении необычных оснований в ДНК сразу начинает синтезироваться измененный белок, а синтез нативного белка прекращается. Хотя эти данные не позволяют еще решить вопрос окончательно, однако они указывают на трудности, возникающие при принятии гипотезы о том, что роль матрицы играет стабильная рибосомная РНК. [11]
Растворимая РНК играет роль адаптера и переносчика аминокислот. На ее долю приходится около 10 % клеточной РНК. Каждая аминокислота имеет, помимо своего собственного специфического активирующего фермента, по меньшей мере одну специфическую s - PHK. В состав s - PHK входит примерно 80 нуклеотидов и среди них некоторое количество необычных оснований. Эти особенности коренным образом отличают растворимые РНК от рибосомной РНК. Необычные основания образуются, вероятно, из обычных уже после синтеза молекулы РНК. По-видимому, содержание спиральных форм в молекулах s - PHK довольно высоко. [12]
Растворимая РНК играет роль адаптера и переносчика аминокислот. На ее долю приходится около 10 % клеточной РНК. Каждая аминокислота имеет, помимо своего собственного специфического активирующего фермента, по меньшей мере одну специфическую s - PHK. В состав s - PHK входит примерно 80 нуклеотидов и среди них некоторое количество необычных оснований. Эти особенности коренным образом отличают растворимые РНК от рибосомной РНК. Необычные основания образуются, вероятно, из обычных уже после синтеза молекулы РНК. По-видимому, содержание спиральных форм в молекулах s - PHK довольно высоко. [13]
Страницы: 1