Адениновое основание
Cтраница 1
Адениновое основание также играет важную роль в гидролизе в результате комплексообразования по атому N-7. Ионы Си ( II) слабо влияют на гидролиз СТР до СОР, когда это дополнительное координирование не может осуществляться. Перенос фосфорила полностью зависит от присутствия ионов цинка и, вероятно, протекает через трехкомпонентный комплекс ( 26), в который входят фенантролин, АТР и ион цинка. [1]
 |
Пары комплементарных нуклеиновых оснований. [2] |
Это означает, что адениновому основанию в одной ( епи будет соответствовать тиминовое в другой цепи. [3]
Гуанозинтрифосфат ( GTP) - еще одно высокоэнергетическое соединение, структура которого аналогична структуре АТР, отличие заключается только в том, что вместо аденинового основания в его состав входит гуаниновое. Хотя GTP находит меньшее использование в биологических системах, чем АТР, тем не менее это соединение участвует в некоторых процессах, требующих затраты энергии, например при синтезе пептидной связи на рибосомах. [4]
Ионы марганца ( II), никеля ( II) и кобальта ( II) связываются со всеми тремя фосфорильными группами АТР, так же как и с атомом N-7 аденинового основания. Эти различия в характере комплексообразования между марганцем ( II) и магнием ( II), как следует из 31Р - ЯМР-спектров, необходимо принимать во внимание при проведении модельных опытов по изучению фермент-субстратных взаимодействий с использованием ионов марганца ( II) в качестве парамагнитного зонда. Механистические заключения, вытекающие из исследований, проведенных с использованием ионов марганца ( II), могут не иметь силу in vivo. [5]
Сущность рН - зависимостей и механизмы этих интересных реакций до конца еще не раскрыты. Повышенная каталитическая активность мягких металлов, например иона двухвалентной меди, которые обладают сильным сродством к азоту, позволяет предположить, что происходит образование хелатной связи с участием атома азота адениновой группы и возникает металлофосфатный комплекс с особой реакционной способностью. Что это взаимодействие с адениновым основанием действительно важно для катализа, свидетельствует тот факт, что гидролиз АТФ, катализируемый ионами двухвалентной меди, протекает примерно в 20 раз быстрее, чем гидролиз Y-фенилпропилтрифосфата, где такое взаимодействие исключено. [6]
Характерная особенность производных аденина - их способность влиять на рост растений. Значение аденина обусловлено тем, что он является одним из компонентов нуклеиновых кислот и многих коферментов. Аденин обладает более выраженными свойствами основания, чем пиримидин, но в то же время адениновое основание слабее имидазола. Поскольку и кислотный характер у аденина выражен сильнее, чем у имидазола, в химических реакциях он может участвовать и как основание, и как кислота. Его биологическая активность возрастает, если к аминогруппе в положении 6 присоединяется слабая кислота. По физиологической активности кинетин относится к соединениям, регулирующим рост и развитие растений. Он стимулирует синтез белков, нуклеиновых кислот и соответственно процесс клеточного деления, кроме того, замедляет старение растений. [7]
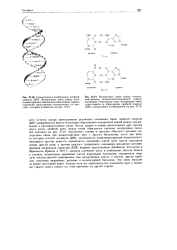
Цепи двойной спирали ДНК удерживаются вместе благодаря образованию водородных связей между основаниями в противоположных цепях. Когда аденин и тимин расположены друг против друга вдоль двойной цепи, между ними образуются прочные водородные связи, как показано на рис. 25.19. Аналогично гуанин и цитозин образуют прочные водородные связи при взаимодействии друг с другом. Вследствие этого две цепи, из которых состоит молекула ДНК, оказываются комплементарными ( взаимодополняющими) - против каждого аденинового основания одной цепи всегда находится тимин другой цепи, а против каждого гуанинового основания находится цитозин. Двойная спиральная структура ДНК, впервые предложенная Джеймсом Уотсоном и Френсисом Криком в 1953 г., сыграла ключевую роль в понимании синтеза белков в клетках, механизмов заражения клеток вирусными частицами, механизмов передачи наследственной информации при размножении клеток и многих других проблем, имеющих важнейшее значение в молекулярной биологии. Эти темы выходят за рамки настоящей книги. [9]
Са, взаимодействующих с фосфатными группами, область титрования полиА смещается к меньшим значениям рН, т.е. спиральная конформация становится менее выгодной. В то же время обработка полиА формальдегидом, благодаря чему аминогруппа аденина оказывается не способной к ионизации, приводит к тому, что молекулы полиА не титруются в области рН 6 и не совершают конформационного перехода в спиральную форму. Видно, что в этом случае, в противоположность тому, что наблюдалось для ДНК и комплексов полиА полиУ - и полиА полиИ, заряд адениновых оснований при переходе спираль-клубок резко падает. [10]
Помимо образования этих различных гетероцепочечных комплексов, некоторые гомополимеры могут в соответствующих условиях самоагрегировать. Изменения констант седиментации, вязкости и светорассеяния коррелируют с кривыми титрования, и эти данные показывают, что при протонировании аденинового остатка образуется относительно жесткий, прерывистый двойной спиральный агрегат из различного числа молекул адениловой кислоты. Так как аминогруппа аденина легко реагирует с формальдегидом, когда полимер находится в беспорядочно скрученном состоянии, но не тогда, когда образован содержащий водородные связи комплекс, вероятно, что агрегация является результатом спаривания адениновых оснований за счет водородных связей. [11]
Это, объясняется тем, что в стабилизации спиральной конформации молекулы полиА существенную роль играет электростатическое взаимодействие положительно заряженных пуриновых оснований с отрицательно заряженными фосфатными группами полинуклеотид-ной цепи. Действительно, при добавлении ионов Са, взаимодействующих с фосфатными группами, область титрования полиА смещается к меньшим значениям рН, т.е. спиральная конформация становится менее выгодной. В то же время обработка полиА формальдегидом, благодаря чему аминогруппа аденина оказывается не способной к ионизации, приводит к тому, что молекулы полиА не титруются в области рН 6 и не совершают конформационного перехода в спиральную форму. Видно, что в этом случае, в противоположность тому, что наблюдалось для ДНК и комплексов полиА полиУ и полиА полиИ, заряд адениновых оснований при переходе спираль-клубок резко падает. [12]
Сравнение энтальпий для ряда олиго-адениловых кислот позволяет сделать очень важный вывод - ДЯ в основном определяется разрушением одной пары вертикально связанных оснований. Величина энтальпии изменяется почти независимо от длины цепи. Так, в 0 01 М LiCl ДЯ АФА равна 10 200 кал / моль, а поли - А 12 450 кал / моль. Исходя из этой модели, авторы рассчитали, что при температуре плавления ( около 45 С) кулоновский сегмент имеет 3 адениновых основания, при 25 С - 7 оснований, а при 10 С - 20 оснований; теплота перехода спираль-клубок для поли - А должна быть около 10000 кал / моль нуклеотида. [13]
Страницы: 1