Глюкозильные остатки
Cтраница 1
Глюкозильные остатки, по-видимому, переносятся только к воде, причем разрыв связей происходит между 1 - м атомом углерода и кислородом. Амилаза содержит кальций в количестве по меньшей мере 1 г-атом на 1 моль. Кальций необходим для ферментативной активности; по-видимому, он способствует сохранению соответствующей конфигурации белка. [1]
Таким образом, основными донорами глюкозильных остатков при биосинтезе целлюлозы являются, по-видимому, гуанозин - и уридиндифосфатглюкоза. [2]
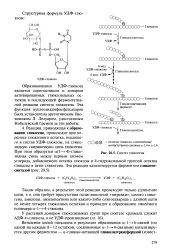 |
Синтез гликогена. [3] |
Образовавшаяся УДФ-глюкоза является переносчиком и донором активированных глюкозильных остатков в последующей ферментативной реакции синтеза гликогена. [4]
Амилазы ( глюкоамилазы, амилоглюкозидазы) - ферменты, отщепляющие глюкозильные остатки последовательно, один за другим, от периферических концов полигликозидных ветвей. [5]
На основании изложенных данных мы пришли к выводу, что в мышцах, как и в печени, содержится трансгликозилаза, катализирующая перенос глюкозильных остатков декстринов. На мальтозу этот фермент действия не оказывает. Таким образом, наши исследования показали, что в мышцах существуют две трансгликози-лазы нефосфоролитического типа, одна из которых действует на мальтозу, другая - на декстрины. [6]
Показано [200], что ксилоглюкан клеточных стенок эндосперма риса состоит из основной цепи p - D - ( l - - 4) - D-глюкана и боковых одноединичных D-ксилозильных или галактозилксилозильных групп, присоединенных к С-6 глюкозильных остатков. Като и др. [170, 171] выделили ксилоглюканы из незрелых злаковых растений ( ячмень, рис и бамбук); предложены их возможные структуры. Хотя общей особенностью таких ксилоглюканов является основная цепь, состоящая из D-глюкозильных остатков, соединенных связью p - D - ( l - 4), и боковых цепей - простых D-ксилозиль-ных групп, присоединенных к С-6 глюкозильных остатков основной цепи, тонкая структура может иметь различия. Содержание D-ксилозы в ксилоглюкане одноклеточных значительно ниже, чем в ксилоглюкане двудольных. Соответствующие большинству фундаментальных, структурных единиц ксилоглюканы двудольных не были получены, хотя фрагменты олигосахаридов из ксилоглюкана однородны. [7]
Рассмотрим некоторые кинетические закономерности ферментативной деградации полимеров на примере действия глюкоами-лазы ( экзо-1 4-а-глюкозидазы или амилоглюкозидазы, КФ 3.2.1.3) из Aspergillus niger. Этот фермент катализирует последовательное отщепление глюкозильных остатков от невосстанавливающего конца мальтодекстринов с уменьшением реакционной способности по мере уменьшения степени полимеризации субстрата. [8]
Фосфорилаза крахмала обнаружена во многих растениях, а из картофеля выделена в кристаллическом виде. Этот фермент катализирует обратимый перенос глюкозильных остатков между глюкозо-1 - фосфатом и невосстанавливающим концом глюкозной цепочки, содержащей сс-1 4-связи. В качестве акцепторов могут действовать различные соединения; однако для фермента, по-видимому, необходима невосстанавливающая цепочка, оканчивающаяся по меньшей мере тремя глюкозильными остатками, соединенными а-1 4-связями, причем замещение в остатках может происходить только у 1-го и 4-го атомов углерода. Наименьшей молекулой, способной действовать в качестве акцептора, является молекула мальтотриозы. Чем больше молекулы, тем они эффективнее; наиболее эффективными оказались соединения с цепочкой, состоящей примерно из 20 глюкозильных остатков, соединенных а-1 4-свя-зями. Фермент действует более активно на разветвленный акцептор, содержащий несколько таких цепочек, чем на акцептор с неразветвленной цепью. Наибольшую активность фермент проявляет по отношению к амилопектину. Фермент не изменяет число невосстанавливающих концевых групп; небольшая молекула акцептора может послужить исходной для образования молекулы больших размеров. Поэтому такой акцептор ( в данной реакции и других ей подобных) называют затравкой. Однако он представляет собой просто второй реагент бимолекулярной реакции. [9]
В числе а-трансглюкозильных ферментов были рассмотрены полисахаридметаболизирующие энзимы: декстрансахараза, амиломальтаза, D-энзим и амилаза Вас. Все они не изменяют а-конфигу-рацию переносимых глюкозильных остатков. [10]
Показано [200], что ксилоглюкан клеточных стенок эндосперма риса состоит из основной цепи p - D - ( l - - 4) - D-глюкана и боковых одноединичных D-ксилозильных или галактозилксилозильных групп, присоединенных к С-6 глюкозильных остатков. Като и др. [170, 171] выделили ксилоглюканы из незрелых злаковых растений ( ячмень, рис и бамбук); предложены их возможные структуры. Хотя общей особенностью таких ксилоглюканов является основная цепь, состоящая из D-глюкозильных остатков, соединенных связью p - D - ( l - 4), и боковых цепей - простых D-ксилозиль-ных групп, присоединенных к С-6 глюкозильных остатков основной цепи, тонкая структура может иметь различия. Содержание D-ксилозы в ксилоглюкане одноклеточных значительно ниже, чем в ксилоглюкане двудольных. Соответствующие большинству фундаментальных, структурных единиц ксилоглюканы двудольных не были получены, хотя фрагменты олигосахаридов из ксилоглюкана однородны. [11]
Это правило заключается в том, что молекулярное вращение гликозида ( Ш ] в [ а ] вМ / 100) приблизительно равно алгебраической сумме молекулярных вращений сахарного остатка и агликона. В случае олигосахаридов за агликон принимаются мрносахаридное звено или звенья, связанные с нередуцирующим концевым остатком. Практически задача определения конфигурации гли-козидной связи сводится к вычислению молекулярного, а отсюда и удельного вращения для гликозидов с а - и fl - связями и сравнению с вычисленными величинами экспериментально найденного [ а ] в исследуемого соединения. За удельное вращение нередуцирующего концевого остатка принимают вращение гликозида соответствующего сахара с оптически неактивным агликоном, например, если в оли-госахариде концевой остаток - D-глюкозильный остаток, - исходными величинами будут удельные вращения метил-а - О-глюкозида и метил-р - О-глюкозида. Отсюда вычисляют молекулярное вращение а - и fl - глюкозильных остатков. [12]
Фосфорилаза крахмала обнаружена во многих растениях, а из картофеля выделена в кристаллическом виде. Этот фермент катализирует обратимый перенос глюкозильных остатков между глюкозо-1 - фосфатом и невосстанавливающим концом глюкозной цепочки, содержащей сс-1 4-связи. В качестве акцепторов могут действовать различные соединения; однако для фермента, по-видимому, необходима невосстанавливающая цепочка, оканчивающаяся по меньшей мере тремя глюкозильными остатками, соединенными а-1 4-связями, причем замещение в остатках может происходить только у 1-го и 4-го атомов углерода. Наименьшей молекулой, способной действовать в качестве акцептора, является молекула мальтотриозы. Чем больше молекулы, тем они эффективнее; наиболее эффективными оказались соединения с цепочкой, состоящей примерно из 20 глюкозильных остатков, соединенных а-1 4-свя-зями. Фермент действует более активно на разветвленный акцептор, содержащий несколько таких цепочек, чем на акцептор с неразветвленной цепью. Наибольшую активность фермент проявляет по отношению к амилопектину. Фермент не изменяет число невосстанавливающих концевых групп; небольшая молекула акцептора может послужить исходной для образования молекулы больших размеров. Поэтому такой акцептор ( в данной реакции и других ей подобных) называют затравкой. Однако он представляет собой просто второй реагент бимолекулярной реакции. [13]
Страницы: 1