Остов - цепь
Cтраница 1
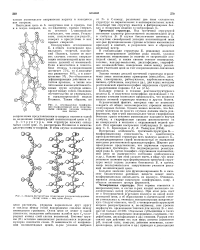 |
Модель - структуры ( пунктиром указаны водородные связи. слева - вид цепи сбоку. [1] |
Остов цепи не лежит в одной плоскости; вследствие небольших изгибов при С - угле-родных атомах слой слегка волнистый. Боковые группы ( R) остатков аминокислот располагаются перпендикулярно плоскости слоев. [2]
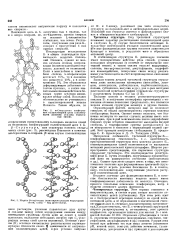 |
Модель fl - структуры ( пунктиром указаны водородные связи, слева - вид цепи сбоку. [3] |
Остов цепи не лежит в одной плоскости; вследствие небольших изгибов при Са-угле-родных атомах слой слегка волнистый. Боковые группы ( R) остатков аминокислот располагаются перпендикулярно плоскости слоев. [4]
Исходя из этого, а также учитывая стерические факторы, препятствующие размещению соседних R-rpynn в полностью развернутой цепочке, Полинг предположил, что остов цепи не лежит строго в одной плоскости, а за счет небольших изгибов при Са-углеродных атомах слой оказывается слегка волнистым, или, как его назвали, складчатым. Остальные периоды ( 4 7 Аи 10 А) при этом не меняются. Было предположено существование двух типов складчатых слоев ( фиг. [5]
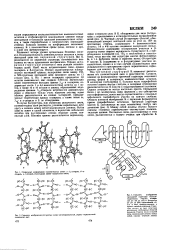 |
Схематич. изображение р-структур. слева-антипараллельный, справа-параллельный складчатый лист.| Схематич. изображение. [6] |
В случае р-структуры, или структуры складчатого листа, полипептидные цепи растянуты, уложены параллельно друг другу и связаны между собой водородными связями. Остов цепи не лежит в одной плоскости, а вследствие небольших изгибов при а-углеродных атомах образует слегка волнистый слой. [7]
Нуклеиновые кислоты представляют собой высокомолекулярные соединения, состоящие из мононуклеотидов, в которых цепи образуются за счет монофосфатных связей между положением 5 одного нуклеозида и положением 3 другого. Остов цепи, таким образом, состоит из фосфатных и сахарных звеньев, к которым через регулярные интервалы присоединены пуриновые и новые основания. [8]
Такая конформация полипептидных цепей предопределяет возможность параллельной и антипараллельной укладки макромолекул и возникновение между ними достаточно прочных межмолекулярных контактов. Остов цепи не лежит в одной плоскости: вследствие изгибов при Са-атомах слой белкового субстрата оказывается слегка волнистым. Боковые заместители аминокислотных звеньев располагаются перпендикулярно плоскости слоев. В фибриллярных белках полипептидные цепи в р-структурах упакованы антипараллельно. [9]
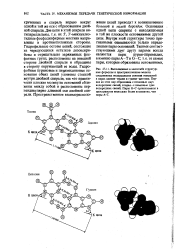
Две цепи в этой спирали антипараллельны, т.е. их 5, З - межнуклео-тидные фосфодиэфирные мостики направлены в противоположные стороны. Гидрофильные остовы цепей, состоящие из чередующихся остатков дезоксири-бозы и отрицательно заряженных фосфатных групп, расположены на внешней стороне двойной спирали и обращены в сторону окружающей ее воды. Гидрофобные пуриновые и пиримидиновые основания обеих цепей уложены стопкой внутри двойной спирали, так что практически плоские молекулы оснований сближены между собой и расположены перпендикулярно длинной оси двойной спирали. [11]
Таким образом, если пептидная цепь полностью выпрямлена и естественно натянута, то скелет ее имеет зигзагообразную форму, приближающуюся к форме зубьев пилы. Боковые цепи ( R) при этом выступают соответственно чередованию пептидных группировок попеременно, то сверху, то снизу плоскости остова цепи ( см. схему на этой стр. Длина такой цепи может быть произвольной. Толщина остова принимается равной 4 5 А. [12]
При наличии противоионов натрия, калия и цезия и при относительной влажности 75 % структура ДНК соответствует А-форме. В такой структуре сохраняется форма правовннтовой двойной спирали, а отличия от предыдущей формы состоят в том, что основания упакованы не горизонтально, а наклонно: при этом меняется шаг спирали и число нуклеотидов на один виток, остов цепи также слегка меняет свою конфигурацию. Очень сходной с данной структурой в растворах обладают двухцепо-чечные РНК и ДНК / РНК-гибриды. Наличие в молекулах РНК лишних 2 -гидрок-сильных групп не позволяет им принимать В-форму. Поэтому в процессе транскрипции РНК с ДНК-матрицы последняя, по-видимому, приобретает конфигурацию А-формы. Различные органические растворители и белок вызывают переход ДНК из В - в А-форму. [13]
Тот факт, что изменения в растворимостях и коэффициентах активности существенны уже при относительно низких концентрациях ( в наиболее благоприятных случаях - 0 1 моль / л), указывает на то, что они вызваны скорее прямым взаимодействием между молекулами, чем неспецифическим влиянием на растворитель. Константы ассоциации, вычисленные из данных по растворимости и коэффициентов активности ( типа приведенных на рис. 1), успешно объясняют зависимость положения сдвигов в спектрах ЯМР от концентрации. Неблагоприятное изменение энтропии ассоциации отдельных молекул в разбавленном растворе значительно меньше для оснований, которые связаны с ковалентным остовом цепи нуклеиновой кислоты. Поэтому почти несомненно, что механизм, который ответствен за взаимодействие отдельных молекул основания, является главным фактором, обеспечивающим устойчивость простой или двойной спирали полинуклеотида в водном растворе. [14]
Страницы: 1