S-частица
Cтраница 1
S-частицы также распадаются на пару дублетов. То же самое относится к К-частицам. Но Л - частица образует синглет, а частицы S, 2 и 2 - - триплет. [1]
Эта возможность естественно связывается с S-частицей. [2]
Отсутствие любого из них сказывается на функции 30 S-частицы. Все белки действуют согласованно, что необходимо и для самосборки. Кинетика самооборки отвечает реакции первого порядка, что означает наличие медленной, лимитируюшей процесс перестройки некоторой промежуточной системы. Сильная зависимость скорости самосборки от температуры показывает, что для этой перестройки требуется свободная энергия около 160 кДж / моль рибосом. Са-мосборка рибосом из всех компонент in vitro происхо-дит примерно за 5 мин. [3]
Кроме столкновении с отклонением на большие углы и образования S-частиц при близких столкновениях, частица, проходящая через вещество, испытывает большое число менее сильных столкновений, совместный эффект которых, как уже упоминалось, вызывает постепенное торможение и рассеяние частиц. Хотя, как мы увидим, явления торможения и рассеяния тесно связаны между собой, будет удобно, для того чтобы сделать статистическое рассмотрение как можно более ясным, сначала рассмотреть более детально проблему торможения. [4]
Роль рРНК специфична, но не абсолютна - функциональные 30 S-частицы получаются из 16 S - pPHK одного вида бактерий и рибосомных 30 S-белков другого вида. [5]
Это своеобразие заключается в том, что возбужденные состояния частиц резко разделяются на два класса: метастабильные состояния, аналогично тем, которые рассматривались выше, как Л0 -, 2 - и S-частицы, с относительно большим временем жизни ( - 10 - 10 сек), и коротко-живущие состояния со временем жизни порядка ядерных. Для последних характерна возможность быстрых переходов из возбужденного состояния нуклона, например, в его основное состояние. [6]
Затем на оси абсцисс откладывают значения толщины стенки счетной трубки и восстанавливают перпендикуляр до пересечения с кривой. Ордината точки пересечения определяет искомую долю S-частиц, могущих проникнуть в счетную трубку. [7]
Обнаружение более тяжелых гиперонов и мезонов представляло бы серьезное по своим последствиям научное событие. Поэтому специальные эксперименты по поискам более тяжелых частиц крайне интересны. Очень желательно посмотреть в специальных камерах большого размера, не существуют ли гипероны больших масс с большим числом каскадов распада, чем у S-частиц. [8]
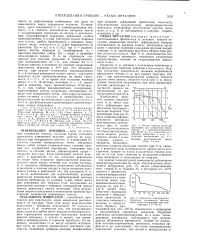 |
Снижение электросопротивления, вызванного облучением меди нейтронами при температуре 17 - 19 К. [9] |
Классификация переходов, обязанных слабым взаимодействиям, в к-рых участвуют только барионы или меаоны, удобно производится с помощью квантового числа странности S, к-рое определяется из равенства: Q / e л / 2 4 - тг - f - S / 2, где п - разность между числом барионов и антибарионов, тг - проекция изотопич. Каждая единица в величине AS отвечает уменьшению вероятности перехода в 1020 раз. S-частица, имеющая S - 2, распадается каскадно, через промежуточную 2-частицу. Этот заряд противоположен но знаку у нейтрино и антинейтрино. [10]
При седиментации рибосом в ультрацентрифуге наблюдаются резкие границы. По аминокислотному составу эти частицы не различаются. При концентрации Mg, равной примерно 10 - 2 М, 70 S-частицы ди-меризуются. Электронные микрофотографии подтверждают выводы, сделанные на основании данных о седиментации и диффузии. [11]
 |
Схема 70S -, 30S - и SOS-частиц. [12] |
Для рибосом митохондрий и хлоропластов эукариотов константа седиментации близка к 70 S, а не к 80 S. S-рибосомы состоят из двух субъединиц - 30 S и 50 S, на которые они диссоциируют при уменьшении концентрации двухвалентных ионов ( Mg) или при увеличении концентрации одновалентных ионов. Форма и размеры 70 S -, 30 S - и 50 S-частиц показаны на рис. 9.11. Аналогичным образом 80 S-рибосомы состоят из 60 S - и 40 S-частиц. [13]
Для рибосом митохондрий и хлоропластов эукариотов константа седиментации близка к 70 S, а не к 80 S. S-рибосомы состоят из двух субъединиц - 30 S и 50 S, на которые они диссоциируют при уменьшении концентрации двухвалентных ионов ( Mg) или при увеличении концентрации одновалентных ионов. Форма и размеры 70 S -, 30 S - и 50 S-частиц показаны на рис. 9.11. Аналогичным образом 80 S-рибосомы состоят из 60 S - и 40 S-частиц. [14]
На SOS-частице находится пептидилтрансферазный центр, обеспечивающий перенос пептидила. Происходит периодическое размыкание и смыкание-рибосомы. На рис. 8.13, в рибосома сомкнута, аминоациль-ный конец комплекса аминоацил-т РНК располагается вплотную к этерифицированному карбоксилу пептидила. Смыкание субъединиц рибосомы ( б - - в) сопровождается образованием пептидной связи. Затем происходит транслокация - тРНК переходит от пептидил-т РНК с 30 S - на 50 S-частицу, увлекая за собой связанный с ней кодон мРНК и вытесняя деапи-лированную тРНК из пептидил-т РНК-связывающего участка. Затем на расположенный в 30 S-частице новый кодон поступает новый комплекс аминоацил-т РНК и цикл начинается сначала. [15]
Страницы: 1 2