Большинство - макромолекул
Cтраница 2
При замещении атомов водорода др. атомами или группами, напр, атомами хлора [ радиус 0 18 нм ( 1 8 А) или фтора [ радиус 0 15 нм ( 1.5 А) ], в большинстве случаев цепь уже не может сохранить плоскую конформацию, поскольку большие атомы вызывают значительные напряжения в макромолекуле. Поэтому большинство полимерных макромолекул имеет спиральную конформацию. В этом случае период идентичности может включать один или неск. В интервале темп-р 20 - 30 С цепь слегка раскручивается, так что на период идентичности приходится пятнадцать звеньев. Форма макромолекулы политетрафторэтилена близка к цилиндрической. При темп-ре выше 30 С структура становится частично беспорядочной; цепи, не нарушая взаимного расположения, согласованно колеблются или вращаются вокруг своих осей. [16]
При замещении атомов водорода др. атомами или группами, напр, атомами хлора [ радиус 0 18 нм ( 1 8 А) ] или фтора [ радиус 0 15 нм ( 1 5 А) ], в большинстве случаев цепь уже не может сохранить плоскую конформацию, поскольку большие атомы вызывают значительные напряжения в макромолекуле. Поэтому большинство полимерных макромолекул имеет спиральную конформацию. В этом случае период идентичности может включать один или неск. В интервале темп-р 20 - 30 С цепь слегка раскручивается, так что на период идентичности приходится пятнадцать звеньев. Форма макромолекулы политетрафторэтилена близка к цилиндрической. При темп-ре выше 30 С структура становится частично беспорядочной; цепи, не нарушая взаимного расположения, согласованно колеблются или вращаются вокруг своих осей. [17]
Газожидкостная хроматография представляет собой очень ценный метод анализа низкомолекулярных соединений, например мономеров, различного вида добавок, растворителей. Трудность использования этого метода для анализа полимеров заключается в том, что большинство макромолекул даже при повышенных температурах имеют слишком низкое давление паров, чтобы проходить через колонки такого типа. Частично эту проблему удается решить путем применения пиролитической газовой хроматографии ( разд. [18]
В реальных молекулах внутреннее вращение всегда в той или иной степени заторможено из-за взаимодействия между валентно не связанными атомами и атомными группами. Как видно из табл. 1 и 2, заторможенность внутреннего вращения в макромолекулах существенным образом сказывается на их размерах и дипольных моментах: средние размеры большинства типичных макромолекул винилового ряда в 1 8 - 2 5 раза больше, а средние дипольные моменты, как правило, меньше, чем для свободного вращения. [19]

Животные клетки осуществляют перенос макромолекул через плазматическую мембрану путем эндоцитоза и экзоцитоза ( см. гл. В клетках растений эти процессы сильно затруднены из-за наличия жесткой клеточной стенки и тургорного давления. Ограниченная проницаемость клеточной стенки не позволяет микрочастицам и большинству макромолекул вступать в прямой контакт с внешней поверхностью плазматической мембраны; поэтому растительные клетки за очень редким исключением не могут поглощать такие частицы путем эндоцитоза. Это ограничение распространяется даже на жидко-фазный эндоцитоз малых молекул ( разд. Тем не менее плазматическая мембрана растительной клетки образует многочисленные окаймленные ямки, которые, как полагают, отшнуровывают-ся, образуя окаймленные эндоцитозные пузырьки ( рис. 19 - 35) ( см. также разд. [21]
Чем выше молекулярная масса, тем выше температура, при которой вязкое течение начинает превалировать над высокоэластичными деформациями, определяемыми молекулярными зацеплениями. Вязкое течение проявляется в том случае, когда в зацеплениях напряжение релаксирует в течение периода времени, соизмеримого с длительностью эксперимента. Поскольку для типичного полимера характерна величина молекулярной массы отрезков цепей между зацеплениями порядка 20 000, большинство макромолекул участвуют в нескольких зацеплениях. Число зацеплений, приходящихся на одну цепь, возрастает с увеличением молекулярной массы. [22]
Пробелы в наших знаниях о белках тесно связаны с тем, что свойства белков зависят не только от химического состава и порядка сочетания аминокислотных остатков в полипептидной цепи, но также и от расположения этих цепей в пространстве. Свойства молекулы, содержащей компактно упакованные длинные пептидные цепи, отличаются от свойств той же молекулы, содержащей пептидные цепи в развернутом виде. В отличие от большинства других фибриллярных макромолекул, пептидные цепи белков могут сохранять свой специфический узор в течение долгого времени. [23]
Такие нити имеют низкую прочность и чрезмерно высокое удлинение. Это объясняется малой упорядоченностью структуры нитей, так как только небольшая часть макромолекул ориентирована параллельно оси нити. Чем больше ориентированы макромолекулы, тем большее их число сопротивляется разрыву одновременно и в равной степени. Поэтому прочность нити может быть значительно увеличена путем перегруппировки и ориентации большинства макромолекул в направлении, параллельном оси нити. Это достигается вытягиванием нити после формования. [24]
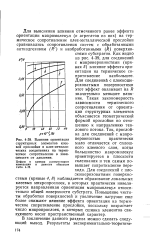 |
Влияние ориентации структурных элементов клеевой прослойки в клее-металли. [25] |
Как видно из рис. 4 - 39, для соединений с макронеровностями ( кривая 5) влияние эффекта ориентации на термическое сопротивление наибольшее. Для соединений с плоскошероховатыми поверхностями этот эффект оказывает на R значительно меньшее влияние. Такая закономерность зависимости термического сопротивления от ориентации структурных элементов объясняется геометрической формой прослойки по отношению к направлению теплового потока. Так, прослойка для соединений с макронеровностью практически мало отличается от сплошного клеевого слоя, когда большинство макромолекул ориентируется в плоскости склеивания и тем самым повышает сопротивление перехода. Для соединений с плоскошероховатыми поверхностями ( кривые 4, 8) наблюдается образование локальных клеевых микропрослоек, в которых практически нивелируется направленная ориентация макромолекул относительно общей поверхности субстрата. Повышение чистоты обработки поверхностей и увеличение нагрузки еще более снижают влияние эффекта ориентации на термическое сопротивление прослойки, поскольку возрастает число микропрослоек, в которых сшивка элементов свя-зурщего носит пространственный характер. [26]
Страницы: 1 2