Перенос - фосфат
Cтраница 1
Перенос фосфата в положение С2 катализируется фосфогли-цератмутазой. В качестве косубстрата участвует 2 3-бисфосфогли-церат и необходимо присутствие ионов Mg2: 2 3 - ФГК - 2 2 - ФГК. [1]
Потенциал переноса фосфата на воду равен - - 29 кДж / моль. Эта энергия может быть перенесена вместе с фосфатной группой и на другой акцептор. Так, для образования аргининфосфата из аргинина и НРС42 - стандартная энергия Гиббса равна 29 кДж / моль. Реакция протекать не может. Но если одновременно происходит гидролиз АТФ, доставляющий энергию, то в целом процесс аргинин АТФ аргинин-фосфат АДФ возможен, так как АС 0 и в смеси исходных веществ идет реакция до установления равновесного состояния. [2]
Можно думать, что перенос фосфата и фосфорилирование связаны скорее с первичной обратимой фиксацией двуокиси углерода ( как в хемосинтезе, так и фотосинтезе), а не с восстановлением углекислого газа до углеводов. Исследования Фоглера и Умбрейта показывают, насколько важно количественное изучение обмена веществ автотрофных бактерий для понимания тесно связанных явлений хемосинтеза и фотосинтеза. [3]
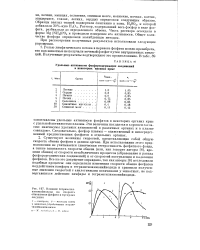 |
Влияние тетрамотил-аммонийиодида на скорость обновления фосфата в мускулах лягушки. [4] |
Ролью лимфатического потока в переносе фосфата можно пренебречь, так как животные не получали меченый фосфат путем внутривенных инъекций. Полученные результаты подтверждают это предположение. [5]
Киназы - ферменты, катализирующие перенос фосфата от АТФ ( или - значительно реже - от другого нуклеозидтрифосфата) к субстрату. [6]
Обращению процесса препятствуют два термодинамически выгодных переноса фосфата от АТР к глюкозе. [7]
Постоянный уровень АТР поддерживается за счет переноса фосфата от креатин-фосфата. [8]
Образуются в результате катализируемого ферментом протеинкиназой переноса фосфата АТФ на гидроксиль-ную группу к-ты в уже сформированной молекуле белка. ФП-фосфа-тазой играют большую роль в гормональной регуляции активности мн. [9]
Синтез нового белка идет параллельно с заметным переносом фосфата из кислотно-нерастворимой фосфатной фракции. [10]
На вопрос о том, протекает ли рассматриваемая реакция переноса фосфата посредством разрыва связи Р - О или X - О, часто можно ответить, анализируя продукты реакции, хотя ферментативные реакции могут быть сложнее, чем представляются при поверхностном рассмотрении, и нередко включают образование промежуточных соединений. Если нуклеофилом служит вода, то расщепление связей Р - О и X - О приводит к одним и тем же продуктам реакции и использование воды, меченной 18О, является единственным способом сделать выбор. [11]
Отсюда можно было сделать вывод, что при таком переносе фосфата не происходит уравновешивания его с неорганическим фосфатом среды, а следовательно, не достигается и лаби-лизация. [12]
Хотя в участии металлов в нейтрализации зарядов при ферментативных реакциях переноса фосфата не приходится сомневаться, можно отметить, что эта функция не обязательно выполняется металлом. Белки содержат и другие заряженные центры, не содержащие металл, - амилные и имидазольные группы, которые с успехом могут участвовать в связывании отрицательно заряженных фосфатных групп. Многие ферменты, в том числе некоторые из переносящих фосфат, работают в отсутствие металлов, хотя их субстраты заряжены отрицательно. [13]
Эти процессы делятся на два типа: окислительно-восстановительные и связанные с переносом фосфатов; реакции первого типа обеспечивают энергию, а второго - механизм для ее переноса. Переносчиком фосфатных ионов является аденозин. [14]
В одной группе реакций используется какой-либо субстрат, обладающий высокой способностью к переносу фосфата. Пируваткиназа у факультативных анаэробов обычно неактивна, особенно в условиях аноксии; поэтому для синтеза высокоэнергетических фосфатов используются только две из этих реакций. [15]
Страницы: 1 2 3