Задержка - дробление
Cтраница 1
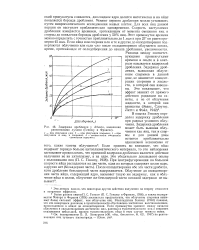 |
Задержка дробления у Arbacia, вызванная рентгеновыми лучами ( Геншоу и Фрэнсис. [1] |
Задержка дробления, вызванная облучением спермиев в данной дозе, не зависит от концентрации спермиев и жидкости, в которой они взвешены. [2]
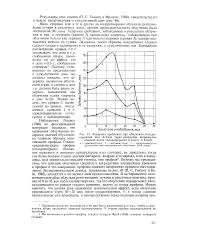 |
Задержка дробления при облучении оплодотворенных яиц Arbacia через различные промежутки времени после оплодотворения ( Геншоу и Коэн. [3] |
Задержка дробления, наблюдаемая в результате облучения и яиц, и спермиев ( кривая 3) превосходит задержку, наблюдаемую при облучении только яиц ( кривая /) или только спермиев ( кривая 2), однако оказывается меньше их суммы. Задержка, таким образом, неаддитивна. Однако следует ожидать аддитивности не задержки, а кумулятивных доз. Можно видеть, что кривая 3, вычисленная из данных, описываемых кривыми 1 и 2, удовлетворительно согласуется с наблюдениями. [4]
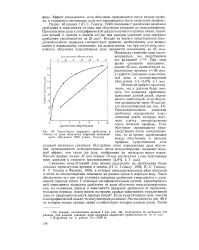 |
Зависимость задержки дробления у Arbacia от дозы облучения спермиев ( интенсивность облучения 7800 р / мин, Геншоу. [5] |
Продолжительность задержки дробления определяется кумулятивной дозой, которую получила клетка непосредственно перед началом профазы. [6]
Яйца были оплодотворены тотчас же после облучения, и задержка дробления оказалась соответственно равной 21 и 12 мин. [7]
Однако у Cumingia, у которой при наличии периода покоя между облучением и оплодотворением восстановление не происходит, задержка дробления, вызванная действием облучения в данной дозе, оказалась не зависящей от интенсивности облучения. [8]
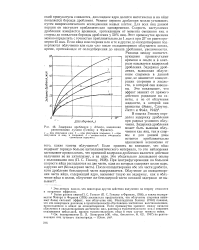 |
Задержка дробления у Arbacia, вызванная рентгеновыми лучами ( Геншоу и Фрэнсис. [9] |
Если принять во внимание, что яйцо содержит гораздо больше цитоплазматического материала, то эти наблюдения заставляют предполагать, что причиной задержки дробления является действие излучения не на цитоплазму, а на ядро. После оплодотворения обе эти части дробятся, хотя дробление безъядерной части задерживается. [10]
Однако, несмотря на хранение при низкой температуре, они не оправляются от другого, также вызванного облучением изменения, которое обусловливает задержку дробления. [11]
Поскольку доза, использованная в этом опыте, не указана, вычислим ее, исходя из того, что 30-минутная экспозиция вызывает задержку дробления, равную 21 мин. Отсюда определим задержку, ожидаемую при 150-минутной экспозиции, которая будет равна 14 мин. Это хорошо согласуется с наблюдавшейся в опыте задержкой дробления, равной 12 мин. [12]
Снижение кумулятивной дозы можно проследить на протяжении более длинных промежутков времени в опытах ( П. С. Геншоу, 1932; П. С. Геншоу, К. Т. Геншоу и Фрэнсис, 1933), в которых неоплодотворенные яйца облучали и затем до оплодотворения помещали на разные сроки в морскую воду. Было обнаружено, что при этих условиях задержка дробления уменьшается с удлинением периода покоя. Если кумулятивную дозу нанести в логарифмической шкале, то получим прямую линию. Это показано на рис. 49, б на котором четыре прямые линии соответствуют четырем разным дозам. [13]
Эффект уменьшается, если облучение производится после начала профазы, и становится ничтожным, если оно производится после окончания профазы. На рис. 48 кривая / ( П. С. Геншоу, 1940) показывает увеличение задержки дробления в зависимости от дозы при облучении спермиев до оплодотворения. Исходя из нашего представления ( продолжительность задержки соответствует времени, необходимому для возвращения к нормальному состоянию), мы делаем вывод, что при отсутствии дальнейшего облучения кумулятивная доза снижается наполовину за 25 мин. [14]
Поскольку доза, использованная в этом опыте, не указана, вычислим ее, исходя из того, что 30-минутная экспозиция вызывает задержку дробления, равную 21 мин. Отсюда определим задержку, ожидаемую при 150-минутной экспозиции, которая будет равна 14 мин. Это хорошо согласуется с наблюдавшейся в опыте задержкой дробления, равной 12 мин. [15]
Страницы: 1 2