Подвижность - метка
Cтраница 1
Подвижность метки отчетливо проявляется в спектрах ЭПР. [1]
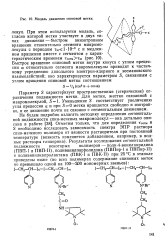 |
Модель движения спиновой метки. [2] |
Параметр S характер изует пространственные ( стерические) ограничения подвижности метки. Уменьшение 5 соответствует увеличению угла прецессии а и при 5 0 метка вращается свободно и изотропно, и ее движение почти не связано с сегментальным движением. [3]
ЭПР сильно иммобилизованных свободных радикалов, а при рН 11 5 подвижность парамагнитной метки сильно увеличивалась в результате разрушения вторичной структуры полинуклеотидов. Наличие изобестических точек указывало на то, что этот переход носит кооперативный характер. При исследовании полицитидиловой кислоты авторы наблюдали наложение двух сигналов ЭПР, из чего следовало, что лишь часть молекул или не все участки молекул этого полинуклеотида имеют упорядоченную вторичную структуру. Полиуридиловая кислота, как оказалось, находится в состоянии статистического клубка даже при значениях рН, близких к нейтральным. [4]
 |
Метка в цепи ( а и изменение характерных времен внутреннего вращения тр ( М ( р для цепи с центральной нагруженной меткой (. м 4. по мере удаления от метки ( б.| Отношение времен TJ ( M. [5] |
Таким образом, последовательное усиление динамической корреляции между движениями звеньев в цепи при ужесточении связей, валентных углов, повышении барьеров внутреннего вращения уменьшает различия в подвижности метки и остальных звеньев цепи. [6]
Очень разбавленный раствор освещают поляризованным УФ-светом, возбуждающим поляризованную же люминесценцию меток. Подвижность меток приводит к деполяризации люминесценции, происходящей по экспоненциальному закону того же вида, что затухание эффекта Керра. Время релаксации поляризованной люминесценции т является прямой мерой подвижности метки, а подвижность эта ( особенно - хотя и не обязательно - если метка находится в главной цепи) однозначно характеризует локальные или общие конформационные степени свободы. [7]
Мы уже ушли от принципа ТВЭ, ибо индикатор ( метка), введенный в среду, характеризует температурную зависимость плотности или ( с некоторыми оговорками) вязкости разных элементов объема этой среды, безотносительно к частоте. Подвижность метки либо есть, либо ее нет. Опыты основаны не на сканировании, как при первоначальном рассмотрении смысла стрелки действия, а на зондировании, притом локальном. [8]
Спин-метка чувствительна к своему окружению, ее спектр ЭПР зависит от конформационного состояния биополимера. Подвижность метки связана с локальной подвижностью биополимера и подвижностью его молекулы как целого. С помощью спин-меток установлены важные особенности строения и динамики белковых молекул. Эти методы весьма эффективны и при изучении биологических мембран. [9]
Сравнивая времена корреляции иминоксильных свободных радикалов, привитых к сульфгидрильным группам расщепленных дисульфид-ных связей у-глобулинов и к SH-группе сывороточного альбумина, обработанного мочевиной ( т 1 09 - 10 J) и диоксаном ( т - 2 04 - 1СГ9), естественно прийти к выводу, что фрагменты полипептидных цепей у-глобулинов, к которым присоединены свободные радикалы, не обладают упорядоченной вторичной структурой. Особенно велика подвижность иминоксильных меток в у-глобулине кролика, где она даже превышает таковую для сывороточного альбумина, денатурированного мочевиной. [10]
Исходя из данных по изучению вторичной и третичной структуры белков оптическими и гидродинамическими методами [47], естественно полагать, что состояния спин-меченого фрагмента полипептидной цепи после обработки белка мочевиной и диоксаном отличаются отсутствием или наличием а-спиральной структуры. Наличие вторичной структуры существенно увеличивает жесткость спин-меченого участка цепи макромолекулы, а потому подвижность парамагнитной метки в присутствии мочевины значительно больше, чем в присутствии диоксана. [11]
Это означает, что при решении обратной задачи совместное определение параметров, ха7 рактеризующих подвижность метки и параметров спин-гамильтониана, некорректно. К этим общим принципам, следующим из теории, необходимо добавить следующее. Так как решение обрат ной задачи итеративным путем невозможно, то остается путь моделирования с использованием спектральных параметров, которые можно измерять в условиях эксперимента. [12]
Очень разбавленный раствор освещают поляризованным УФ-светом, возбуждающим поляризованную же люминесценцию меток. Подвижность меток приводит к деполяризации люминесценции, происходящей по экспоненциальному закону того же вида, что затухание эффекта Керра. Время релаксации поляризованной люминесценции т является прямой мерой подвижности метки, а подвижность эта ( особенно - хотя и не обязательно - если метка находится в главной цепи) однозначно характеризует локальные или общие конформационные степени свободы. [13]
Исследование спин-меченого полистирола в растворе показало 85, что времена корреляции спиновой метки близки к тс, определенным из времени спин-решеточной релаксации мета - и пара-протонов полистирола. Значение энергии активации вращения метки составляет 4 3 ккал молъ и близко к величине 4 8 ккал / моль, полученной по данным метода диэлектрической релаксации. Результаты этого сопоставления позволили авторам сделать вывод о том, что подвижность парамагнитной метки в растворе полимера определяется частотой движения сегментов макромолекулы. [14]
Говоря о митохондриальных мембранах, необходимо прежде всего экспериментально установить наличие в них конформа-ционных превращений. В § 3.8 уже были приведены некоторые данные такого характера, относящиеся к возбудимым мембранам. С помощью спиновых меток была изучена сорбция воды мембранами лио-филизованных СМЧ ( а также хлоропластов, см. гл. Время корреляции т, определенное из спектра ЭПР и характезирую-щее подвижность метки, оказалось немонотонно зависящим от содержания сорбированной воды - с его увеличением т проходит вначале через минимум, затем достигает максимума, после чего монотонно убывает. [15]
Страницы: 1