Поиск - гомология
Cтраница 1
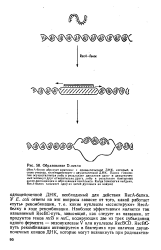 |
Образование D-петли. [1] |
Поиск гомологии осуществляется либо в результате движения одно - и двуцепочеч-ных молекул друг относительно друга, либо в результате повторных актов диссоциации и образования комплекса. [2]
Как же обеспечивается взаимный поиск гомологии между ре-комбинирующими молекулами ДНК. [3]
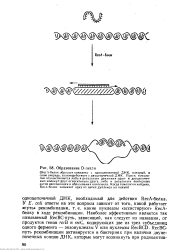 |
Образование D-петли. [4] |
ДНК, который, в свою очередь, взаимодействует с двуцепочечной ДНК Поиск гомологии осуществляется либо в результате движения одно - и двуцепочеч-ных молекул друг относительно друга, либо в результате повторных актов диссоциации и образования комплекса. [5]
 |
Генная конверсия ( о и один из возможных механизмов генной конверсии ( б. [6] |
Мы видим, что по крайней мере у бактерий в общих чертах понятен механизм центрального события гомологичной рекомбинации поиска гомологии и образования полухиазмы. [7]
 |
Генная конверсия ( а и один из возможных механизмов генной конверсии ( б. [8] |
Мы видим, что по крайней мере у бактерий в общих чертах понятен механизм центрального события гомологичной рекомбинации поиска гомологии и образования полу хиазмы. [9]
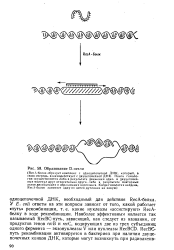 |
Образование D-петли. [10] |
ДНК, который, в свою очередь, взаимодействует с двуцепочечной ДНК. Поиск гомологии осуществляется либо в результате движения одно - и двуцепочеч-ных молекул друг относительно друга, либо в результате повторных актов диссоциации и образования комплекса. [11]
Частичное секвенирование кДНК - клонов и обработка полученных данных полностью автоматизированы. Каждую новую EST сравнивают с теми, которые уже были секвенированы, и если последовательность действительно является новой, ее вносят в базу данных по EST. Для поиска гомологии EST с известными генами или семействами генов и для определения категории, к которой относится функция представляемого ей гена, проводят дополнительные сравнения. К 1995 г. было идентифицировано примерно 300 000 EST из 300 кДНК - библиотек 37 органов и тканей. Примерно 90 000 EST представляют собой различающиеся экспрессирующиеся последовательности человека, из них примерно 10 000 соответствуют генам, роль которых в клетке известна, а остальные 80 000 - еще неоткрытым генам. [12]

У этого организма практически все виды гомологичной рекомбинации зависят от функционирования гена reck ( этот ген играет важную роль и в репарации повреждений ДНК; см. гл. Оказалось, что RecA-белок ( молекулярная масса около 40 кД) является ДНК-зависимой АТРазой, способной заменять одну из цепей дуплекса ДНК на гомологичную. Процесс поиска является активным и требует гидролиза АТР. Показано, что дуплекс ДНК в комплексе с RecA-белком и одноцепочечной ДНК заметно раскручен ( вне зависимости от того, гомологична однонитевая ДНК дуплексу или нет); по всей вероятности, это облегчает поиск гомологии. Другими словами, RecA-белок активно проводит однонитевую миграцию ветви. [14]
У этого организма практически все виды гомологичной рекомбинации зависят от функционирования гена гее А. Оказалось, что RecA-белок ( молекулярная масса около 40 кД) является ДНК-зависимой АТРазой, способной заменять одну из цепей дуплекса ДНК на гомологичную. Процесс поиска является активным и требует гидролиза АТР. Показано, что дуплекс ДНК в комплексе с RecA-белком и одноцепочечной ДНК заметно раскручен ( вне зависимости от того, гомологична однонитевая ДНК дуплексу или нет); по всей вероятности, это облегчает поиск гомологии. [15]
Страницы: 1 2