Реальная популяция
Cтраница 1
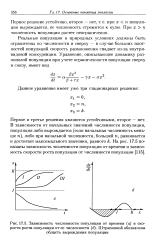 |
Зависимость численности популяции от времени ( а и скорости роста популяции от ее численности ( б. Штриховкой обозначена. [1] |
Реальные популяции в природных условиях должны быть ограничены по численности и сверху - в случае больших плотностей популяций скорость размножения спадает из-за внутривидовой конкуренции. [2]
Рассмотрим теперь реальную популяцию, в которой одни возрастные процессы основаны на закономерностях, рассмотренных в первом из описанных примеров, а другие - во втором; кроме того, имеют место процессы, темп которых зависит от наследственных и средовых различий. Очевидно, наследуемость БВ будет определяться тем, какой тип возрастных изменений избран для его моделирования. В том случае, если БВ отражает по преимуществу общевидовые механизмы старения и мало чувствителен к их индивидуальным модификациям, показатель наследуемости ИБВ будет близок к нулю. На наш взгляд, примерно такая ситуация складывается при моделировании БВ с помощью регрессии клинико-физиологических параметров на КВ. [3]
В реальных популяциях за период развития поколения происходит значительное снижение численности и смертность лишь незначительно ниже или выше рождаемости. [4]
 |
Возрастные периоды состояния у семенных растеши. [5] |
В реальных популяциях границы календарного возраста двух смежных возрастных состояний в какой-то степени перекрываются. Это определяется тем, что индивидуальный ход онтогенеза у отдельных организмов не совпадает и конкретные особи могут достигать определенного возрастного состояния в разные календарные сроки. Функционально же особи данного возрастного состояния сходны. В целом в зависимости от возрастного состава ( возрастного спектра) популяция характеризуется определенными свойствами, отражающими ее онтогенетическое состояние как целостной системы. [6]
Для характеристики динамики реальных популяций используют таблицы выживания, в которых отражают поэтапную выживаемость и смертность особей с указанием или без указания причин смертности. [7]
В настоящее время генетикам и селекционерам известны следующие помехи, существующие в реальных популяциях и так или иначе влияющие на генетико-статистические параметры популяций. [8]
Присущее взаимодействие типа хищник-жертва свойство порождать взаимосвязанные колебания численности вызывало иногда в прошлом надежды на то, что такие же колебания будут обнаружены в реальных популяциях. Эти надежды, однако, следует сразу же умерить по двум соображениям. Во-первых, существует много важных аспектов экологии хищника и жертвы, которые в разработанных моделях пока не учтены; однако, как мы увидим в следующих разделах, эти аспекты могут существенно изменить любые прогнозы. Во-вторых, даже если в популяции наблюдаются регулярные колебания численности, то это совсем не обязательно служит подтверждением модели Лотки-Вольтерры, логистической модели с запаздыванием или любой другой простой модели. Если численность популяции растительноядного животного колеблется, то это может отражать ее взаимодействия с пищевыми объектами или с хищниками. И если численность популяции жертвы циклически меняется в силу присущих ей свойств, то численность хищников может повторять эти циклы, даже если само взаимодействие хищник-жертва их не вызывает. Таким образом, взаимодействия типа хищник-жертва могут порождать регулярные колебания численности обеих взаимодействующих популяций и могут усиливать такие колебания, если они существуют вследствие других причин; но выявление причины регулярных циклических колебаний в естественных условиях представляет собой, как правило, трудную задачу ( см. разд. [9]
Линейная зависимость между увеличением смертности и плотностью, как указывалось в сноске в главе 3, подвергалась критике ( например, [27]), поскольку линейная зависимость якобы не доказана и не может надлежащим образом отражать рост реальной популяции. Смит, используя эту линейную зависимость, исходил из логистической формулы Ферхюль-ста - Пирла. Безусловно, в специфических случаях нельзя рассчитывать, что рождаемость или смертность будут изменяться в точной линейной зависимости от плотности популяции. Однако линейная зависимость в том виде, как она показана на рисунке 5, является наиболее простым символическим предположением в самом общем виде, отображающим все возможные случаи. В определениях Смита [1849, 1855] для повышенной смертности при увеличении плотности нет указаний на такую линейную зависимость, и его основные положения не связаны с ней. [10]
По сравнению с этим результатом первоначальное уравнение без запаздывания дает монотонное замедление роста численности при всех значениях R. Запаздывание по времени приводит к колебаниям численности в модели и, как можно предположить, оказывает сходное дестабилизирующее влияние на реальные популяции ( см. также разд. [11]
 |
Рост модельной популяции бактерий.| Типичная кривая роста популяции бактерий.| Культура бактерий при 30 С. [12] |
Кривая, полученная на основе данных строки А ( табл. 2.4.), называется логарифмической или экспоненциальной кривой. Тогда в идеальных условиях рост бактерий теоретически должен быть экспоненциальным. Сравним эту математическую модель с кривой роста реальной популяции бактерий, изображенной на рис. 2.15. Отчетливо видны четыре фазы роста. [13]
Таким образом, если исходить из тех основных закономерностей, которые характеризуют поведение отдельного микроорганизма, то следует вывод, что численность популяции должна увеличиваться по экспоненциальному закону, неограниченно стремясь в область высоких концентраций. Несложные расчеты, результаты которых часто встречаются в литературе, дают фантастические количества биомассы потомков одной клетки, образующейся за короткий промежуток времени. Хотя это и не учитывается в приведенной зависимости, естественно предположить, что развитие популяции ограничено ресурсами питательной среды. Исчерпание этих ресурсов приводит к прекращению дальнейшего размножения микроорганизмов, поведение которых в последующие интервалы времени определяется разнообразными закономерностями их выживания и инактивации. Это демонстрирует ограничения приема перенесения закономерностей низшего уровня ( клеточного) на высший ( популяционный): поведение реальной популяции микроорганизмов не соответствует рассмотренным выше экстраполированным закономерностям роста отдельного микроорганизма. [14]
Страницы: 1