Микробная популяция
Cтраница 2
В методе оценки кинетики потребления дополнительного субстрата микробной популяцией для вычисления гетеротрофной активности первоначально не учитывается возможная потеря меченого углерода вследствие образования двуокиси углерода в течение периодов инкубации. [16]
Микробное ограничение производительности означает, что если достигнута микробная популяция максимального размера в условиях, обеспечиваемых данным оборудованием компостирования, нагрузка больше не может увеличиваться в ущерб качеству процесса. Нагрузка сверх максимально допустимой подавит процесс частично или полностью и по меньшей мере приведет только к частичной переработке твердых отходов. [17]
Принципиальным отличием подхода Хиншельвуда к рассмотрению кинетики роста микробной популяции является развитие концепции определяющего этапа цепи метаболических процессов. Распространенные представления об узком месте как звене, в котором реакция протекает с наименьшей скоростью и тем самым определяет кинетику всего процесса в целом, являются справедливыми для линейных последовательных реакций. Когда процесс в целом определяется протеканием реакций, соединенных в циклы и образующих пространственную сетку последовательных переходов, предполагающих альтернативные пути метаболизма в зависимости от конкретных условий, Хиншельвуд, развивая концепцию узкого места, предлагает принцип наибольшей скорости реакции. Суть этого принципа заключается в том, что при наличии различных маршрутов реакций основное значение в общем процессе метаболизма приобретает тот путь, по которому реакция может развиваться при данных условиях с наибольшей скоростью. Любое изменение условий роста приводит не к изменению локальной стадии микробиологического синтеза, а к перераспределению кинетических параметров всей системы. Ограничение общей скорости процесса в сетке химических реакций внутриклеточного метаболизма не обязательно определяется наиболее медленной стадией, а зависит от соотношения констант скоростей ряда отдельных реакций. При этом соотношение ферментов различных этапов процесса микробиологического синтеза, их разрушение, расход, образование и диффузия определяют поведение популяции в целом. [18]
Однако существует немало инфекций, когда основная масса микробной популяции сосредотачивается внутри организма, и оказаться снаружи микробы могут только после повреждения его структур. Но это - дело случая, рассчитывать на него не приходится, нужно искать выход. И вот что происходит в природе. [19]
Кроме определения конечного акцептора водорода, метаболическая активность смешанных микробных популяций может быть оценена по скорости любой другой реакции транспортной электронной системы. [20]
Определение активности специфических ферментов также отражает метаболический потенциал микробной популяции данной экологической ниши. Одна часть таких активностей отражает эн-зиматический потенциал всего сообщества ( активности суммарных дегидрогеназ, эстераз, фосфатаз), другая ( целлюлазная, хи-тиназная, нитрогеназная, денитрифицирующая) - только специфической части сообщества, позволяя, однако, оценить тот или иной его потенциал. Ферменты микроорганизмов, включенных в биогеохимический цикл элементов, важны для изучения микробными экологами. Особенно важны ферменты, вовлеченные в цикл углерода и азота, так как они позволяют оценить устойчивость сообществ и экосистем и их потенциал в отношении минерализации органического вещества. При определении ферментативного потенциала популяций, наряду с перечисленным, измеряют активности липаз, целлюлаз, протеаз, амилаз. При измерении ферментативных активностей in situ важно не нарушать микроокружение данной ниши и следить за температурой, рН, влажностью, Eh, а периоды инкубации ( измерений) должны быть достаточно короткими, чтобы за это время количество микрорганизмов не могло значительно измениться. [21]
Биодеградация нефти при низкой температуре происходит эффективнее при использовании микробных популяций, выделенных при той же температуре. [22]
Изучение видового состава биоценозов имеет значение для составления карт микробных популяций активных илов и выделения форм, которые необходимо адаптировать для очистки нефтесодер-жащих сточных вод. Состав биоценозов не исчерпывается двумя десятками видов. [23]
Была отмечена, таким образом, прямая связь между размерами нативной микробной популяции и скоростью инактивации вирусов. Осталось, однако, неясным, вызвана ли инактивация вирусов токсичными продуктами обмена микроорганизмов или противовирусными энзимами. [24]
Просты и удобны экспериментальные методы определения фитотоксичности: метод проростков и метод угнетения микробных популяций по НА. Красильникову ( методики описаны на стр. [25]
 |
Кривые роста микробной популяции Achromobacter coadunatum на низших алифатических спиртах. [26] |
По мере возрастания молекулярного веса высших алифатических спиртов и уменьшения их растворимости лагфаза роста микробной популяции этой культуры увеличивается. По мере адаптации микробной популяции увеличение лаг-фазы роста приобретает генетическое значение. [27]
Важной особенностью субстрата при компостировании является необходимость содержания в нем веществ, требующихся для жизнедеятельности данной микробной популяции. Таким образом, угле-ро д в необработанных отходах будет отсутствовать до тех пор, пока не появится микробная популяция, способная разрушать угле-родсодержащие вещества. Следовательно, если первые члены микробной сукцессии1 опущены, процесс компостирования будет приостановлен. Однако это происходит очень редко, так как отходы несут в себе большое количество разнообразных микроорганизмов. Важным моментом является то, что чем проще форма, в которой находятся отходы, тем больше это разнообразие и, следовательно, тем быстрее образуется компост. [28]
Происхождение первого ограничения заложено в самом определении процесса компостирования: для его осуществления необходимы организмы, причем определенная микробная популяция. [29]
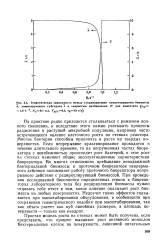 |
Теоретическая зависимость между стационарными концентрациями биомассы х, лимитирующего субстрата с и скоростью разбавления D для хемостата ( мт 1 0 ч 1, Кс0 1 г / л, Ух / с0 5, С0 10 г / л. [30] |
Страницы: 1 2 3 4