Клеточная вакуоль
Cтраница 1
Клеточная вакуоль ограничена белково-лшюидной мембраной - тоно-пластом, наличием которого и объясняется несмешиваемость цитоплазмы и вакуолярного сока и поддерживается определенное осмотическое давление в клетке Срис. [1]
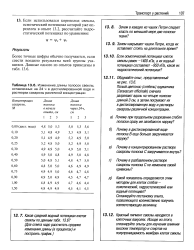 |
Изменение длины полосок свеклы, оставленных на 24 ч в дистиллированной воде и растворах сахарозы различной концентрации. [2] |
Красный пигмент свеклы находится в клеточных вакуолях. Исходя из этого, спланируйте опыты для изучения влияния высоких температур и спиртов на полупроницаемость мембран клеток свеклы. [3]
Ингибиторы амилаз у танидоносных растений ( таннины) находятся в растворенном состоянии в клеточных вакуолях. С помощью гистохимических реакций было установлено, что большинство таннинов локализовано в листьях в обкладочных клетках, окружающих жилки. Это дало основание предположить, что таннины образуются в листьях, откуда проникают в другие части растений. [4]
 |
Участок мякоти яблока, пора - [ IMAGE ] Участок яблока с полностью. [5] |
Нарушение целостности клетки резко меняет внутриклеточное давление, что приводит к разрыву тонопласта, окружающего клеточную вакуоль, и выходу фенольных веществ из вакуоли в цитоплазму. [6]
Плоды не обладают устойчивостью к грибам, вызывающим плодовые гнили. Изучение влияния фенольных соединений на активность пектоди-тических ферментов в модельных опытах показало нам, что эти вещества могут заметно инактивировать ферменты и приостанавливать продвижение гриба по тканям. Гистохимически показана возможность контакта гифы гриба с фенольными веществами, находящимися в вакуоли, в момент разрыва тонопласта, окружающего клеточную вакуоль. Происходит ли в пораженной грибом клетке ингибирование ферментов, подобное тому, которое наблюдается в модельном опыте, или механизм взаимодействия между клеткой и патогеном иной, должны показать дальнейшие исследования. [7]
При изменении нормального физико-химического состояния протоплазмы изменяется отношение ее. Некоторые красители ( нейтральный красный, метиленовый синий, тионин и др.), проникнув в нормальную клетку, оставляют цитоплазму и ядро неокрашенными. В животной клетке они откладываются в цитоплазме в виде мелких гранул или же связываются с некоторыми клеточными включениями. В растительной клетке эти красители гранул не образуют и обычно накапливаются в клеточной вакуоли. Цитоплазма и ядро и здесь остаются бесцветными. При повреждении любой клетки распределение в ней красителя нарушается. Прежде всего парализуется способность клетки откладывать краситель в цитоплазме в виде гранул. Одновременно цитоплазма и ядро приобретают сродство к красителю и начинают все более и более прокрашиваться. [8]
Не исключена возможность, что цинк оказывает влияние на содержание ингибиторов роста растений. К числу таковых, как известно, относятся некоторые фенолкарбоновые кислоты, фла-воноиды и ряд других фенольных соединений. В связи с этим следует упомянуть об исследованиях Рида и Дюфренуа ( Reed, Dufrenoy, 1942), проведенных на грецком орехе и абрикосе. Шаровидные скопления в клеточных вакуолях нуждающихся в цинке растений, формируются в ходе процесса, который авторы считают коацервацией. Частицы дисперсной фазы коллоидов, находящиеся в вакуолях, собираются в шаровидные скопления. Присутствие катехина в центре коацервата обнаруживается по коричневой окраске, появляющейся после окисления, а также выборочным поглощением нейтрального красного. Интересно отметить, что Скугом ( Skoog, 1940) было также открыто окисление катехина в экстракте растений, нуждающихся в цинке. [9]
Страницы: 1