Препарат - нуклеиновая кислота
Cтраница 1
Препараты нуклеиновых кислот первоначально получали путем экстрагирования щелочами с последующим осаждением кислотами. Под воздействием кислот и щелочей макромолекулы нуклеиновых кислот, однако, меняются и их растворы при рН 5 6 и 10 9 [256] теряют характерную для них высокую вязкость, что обусловлено, вероятно, дезагрегацией макромолекул. Оба фермента были выделены Кунитцом из поджелудочной железы в кристаллической форме ( см. гл. [1]
Препараты природных макромолекулярных нуклеиновых кислот получают обычно из нуклеопротеидов, экстрагируемых из тканей солевыми растворами или водой. Если эти нуклеопро-теиды осторожно диализовать или подвергнуть встряхиванию с хлороформом [259], то они расщепляются на белок и полимерную нуклеиновую кислоту. [2]
 |
Изменение нуклеиновых кислот головного мозга при локальном облучении in vivo дозой 2000 р ( сопоставление с ДНК костного мозга. [3] |
На рис. 7 - 44 приведены результаты электронномикроскопи-ческого исследования препаратов нуклеиновых кислот костного мозга. [4]
Биуретовая реакция и проба с аргинином по Веберу [96] служат для обнаружения пептидов в препаратах нуклеиновых кислот. Две другие цветные реакции - предложенная Дише проба на дезоксирибонуклеиновую кислоту и флороглюциновая проба по Эйлеру и Хану - позволяют определить рядом оба вида нуклеиновых кислот. [5]
На колонках с МАК фракции РНК четко отделяются от ДНК, поэтому на колонку можно наносить препараты суммарных нуклеиновых кислот и успешно разделять их. Однако в том случае, когда необходимо расфракционировать большие количества РНК, рекомендуется предварительно очистить ее от ДНК, поскольку присутствие заметных количеств ДНК может привести к замедлению скорости протекания элюента и образованию каналов в колонке, что будет плохо сказываться на разделении фракций РНК. [6]
Как константы седиментации, так и константы диффузии варьируют с изменением концентрации, и если принять во внимание несомненную гетерогенность [163, 164] многих препаратов нуклеиновых кислот, то точное реальное значение их молекулярного веса удается определить довольно редко. Типичными величинами молекулярного веса дезоксирибонуклеиновых кислот являются 4 - 10е - 8 - 10е, хотя в отдельных случаях были получены значительно более высокие значения. [7]
Рибонуклеиновая кислота, выделенная мягкими методами из вируса табачной мозаики ( ВТМ) 1319, 320 ], изучена очень широко, поскольку она является одним из немногих препаратов нуклеиновых кислот, который с полным основанием можно считать гомогенным. Инфекционность таких препаратов и легкая потеря ими инфекционное являются хорошим критерием целостности и других свойств РНК по сравнению с ее свойствами в интактном вирусе. [8]
Однако правильное понимание основного принципа построения нуклеиновых кислот и способа соединения мономерных единиц между собой ( см. формулу V) было достигнуто уже в начале пятидесятых годов, когда исследователи имели в своем распоряжении весьма деградированные препараты нуклеиновых кислот. [9]
Самым старым и самым прямым методом исследования биополимеров следует считать, по-видимому, электронно-микроскопический метод. Недавно Клейншмидт обнаружил, что если препарат нуклеиновой кислоты, непосредственно выделенный из живого организма методом осмотического шока или взятый после предварительного выделения, комплексировать с каким-нибудь основным белком, например с цитохромом с, то его можно нанести на поверхность воды в виде мономолекулярной пленки. ДНК, полученная Томасом и Мак-Хатти при пом: ощи этого метода. Снимок позволяет оценить размеры молекулы ДНК и ясно показывает, что эта молекула замкнута в кольцо. Определенная таким способом длина молекулы, равно как и ее диаметр, соответствуют величинам, которых следует ожидать на основе гипотезы Уотсона - Крика. Показано, что молекулы ДНК из многих источников имеют форму кольца или замкнутой петли. У ДНК фагов Я и Т2 переход от линейной формы к кольцевой осуществляется легко и обратимо. [10]
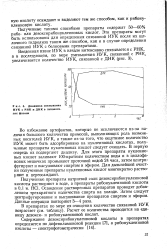 |
Динамика связывания ИУК с РНК и ДНК в гипокоти-лях фасоли. [11] |
Во избежание артефактов, которые не исключаются из-за наличия большого количества примесей, выполняющих роль возможных носителей ИУК, а также из-за того, что ощутимое количество ИУК может быть адсорбировано на нуклеиновых кислотах, полученные препараты нуклеиновых кислот следует очищать. В первую очередь их подвергают диализу. Для этого препараты нуклеиновых кислот заливают 100-кратным количеством воды и в целлофановых мешочках промывают проточной водой 24 часа, затем центрифугируют и высушивают спиртом и эфиром. Для дальнейшей очистки полученные препараты нуклеиновых кислот многократно растворяют и переосаждают. [12]
Определение последовательности оснований необходимо начать с выбора нуклеиновой кислоты и методов определения. Очень часто оба выбора взаимосвязаны, так как один метод может быть пригодным только для небольших молекул, другой окажется единственным эффективным методом в случае нуклеиновых кислот большего размера или отличающихся какими-либо структурными особенностями. Разумеется, пригодность препарата нуклеиновой кислоты во многом зависит от его чистоты. Желательно и фактически необходимо иметь молекулярно-гомогенную нуклеиновую кислоту с минимальным содержанием продуктов разрыва цепи. Уже небольшое количество ( 5 - 10 %) таких примесей может препятствовать однозначному определению последовательности. Даже в случаях, когда требования к молекулярной гомогенности не столь высоки, переход от неоднозначных результатов к правильной структуре требует тем меньше экспериментов, чем более молекулярно-гомогенна нуклеиновая кислота. Рекомендуется также располагать препаратом, не содержащим примесей ненуклеотидного характера, таких, как нукле - азы и другие белки, которые могут мешать анализу и интерпретации полученных данных. [13]
Нуклеиновые кислоты составляют основу растительного и животного мира. Они регулируют синтез белка и с их функцией связаны наследственные свойства организмов. Черноруцким [1911] было установлено, что препараты нуклеиновых кислот увеличивают число лейкоцитов в периферической крови ( нейтрофилов), повышают обмен веществ, стимулируют эритропоэз и фагоцитоз, увеличивая естественные защитные свойства организмов. Значение пири-мидиновых нуклеотидов как компонентов и структурных единиц нуклеиновых кислот было установлено много лет назад. Еще десятилетия назад [ Лазарев, 1947 ] указывалось, что некоторые производные пиримидинов могут применяться в качестве неспецифических стимуляторов общей резистентности организма. Под влиянием аналогов урацила ( 4-метил - 5-оксиметилурацила и 4-мети-лурацила) происходит активация синтеза РНК в печени крыс. [14]
Поэтому методы гель-фильтрации и ионообменной хроматографии не получили широкого распространения при фракционировании нуклеиновых кислот и значительно уступают ультрацентрифугированию и электро-форетическому разделению в геле агарозы, полиакриламидиом геле или их смеси. Поскольку величина отрицательного заряда нуклеиновых кислот и продуктов их расщепления мало зависит от рН, а отношение заряда к молекулярной массе сохраняется практически неизменным, разделение нуклеиновых кислот при электрофорезе определяется не их зарядом, а размером молекул. При наличии маркеров с известной молекулярной массой возможно определение молекулярной массы препаратов нуклеиновых кислот и их фрагментов. [15]
Страницы: 1 2