Приспособленность - вид
Cтраница 1
Наследственная приспособленность видов к определенным условиям среды связана с дифференциацией различных эко-типов в пределах вида. В гл, XII мы уже касались вопроса о том, как отличать экотипы, принадлежащие к одному виду, от разных видов, имеющих разные адаптивные признаки. Основной критерий для решения этой проблемы состоит в том, что экотипы, принадлежащие к одному виду, легко скрещиваются между собой и дают плодовитых гибридов, обладающих нормальной жизнеспособностью; скрещивания же между разными видами, напротив, в большинстве случаев дают худшие результаты. Эти трудности межвидовой гибридизации связаны с несколькими различными явлениями, к рассмотрению которых мы сейчас перейдем. [1]
Дарвина о естественном и искусственном отборах, объяснившие приспособленность видов дикой природы к различным местообитаниям и утерю этих признаков культурными растениями и домашними животными. [2]
Процесс старения следует рассматривать в неразрывной связи со всеми жизненными функциями, определяющими приспособленность вида. Конечным показателем надежности автоматов, конкурирующих за ограниченные ресурсы и самовоспроизводящихся в агрессивной среде, является надежность самовоспроизведения; этот тезис приложим и к биосистемам. [3]
Неравномерность распространения животных в ландшафтах по видовому составу и численности определяется сложными трофическими связями и узкой специализацией и приспособленностью видов. [4]
ИЗМЕНЧИВОСТЬ, разнообразие признаков и свойств у особей и групп особей любой степени родства; один из важнейших факторов эволюции, к-рый обеспечивает приспособленность видов и популяций к изменяющимся условиям существования. Присуща всем живым организмам. [5]
 |
Схема биогеоценоза. [6] |
Все компоненты любого биогеоценоза тесно связаны между собой единством территории, общим потоком энергии ( от Солнца к автотрофам и от них - к гетеротрофам), обменом биогенных химических элементов, сезонными колебаниями климатических условий, численностью и взаимной приспособленностью видов всех уровней организации. [7]
В чем же, однако, усмотрели Паултон и другие авторы идею естественного отбора во втором издании труда Причарда. Изложив приведенные выше соображения о приспособленности видов к условиям обитания, которая, по мнению Причарда, выражает, как мы видели, целесообразное распределение творческой силой созданных ею животных в соответствующих их строению условиях обитания, Причард задает вопрос: Но почему мы должны предполагать, что действие этого закона, приспособления остановилось здесь. Не возможно ли, что разновидности, возникающие в пределах отдельных видов, представляют собой дальнейшие приспособления строения к условиям, в которых трибе предназначено существовать. Разновидности ответвляются от общей формы вида, точно так же как формы вида отклоняются от общего типа рода. [8]
Зависимость фотосинтеза от температуры характеризуется кривой, на которой выделяются точки ( зоны) минимума, оптимума и максимума. Минимальная температура, при которой возможен фотосинтез, видоспецифична и отражает приспособленность вида к температурным условиям среды. [9]
Тем не менее в определенных условиях формирование программы адаптивного саморазрушения возможно. Не менее эффективным, с точки зрения приспособленности вида, может быть увеличение плодовитости в младших возрастах за счет ее снижения в старших. Естественно, что отбор закрепляет комбинированные и сбалансированные решения обоих типов, что не исключает более важной роли одного из них для того или иного вида. [10]
При наследовании возможны мутации из-за радиоактивности или других влияний, в результате которых могут измениться некоторые гены в половых клетках одного из родителей. Измененные гены передаются потомку и придают ему новые свойства. Если эти новые свойства полезны, они скорее всего сохранятся в данном виде, при этом произойдет скачкообразное повышение приспособленности вида. [11]
Шестой и седьмой принципы. Среди гипотез, объясняющих возрастное нарастание смертности, чаще других дискутируется и имеет наибольший практический интерес гипотеза запрограммированного старения, согласно которой на определенном этапе онтогенеза включаются генетические механизмы активного саморазрушения организма, ведущие к старению и смерти. Подчеркнем: речь идет не о том, что старение так или иначе связано с генетической информацией ( это очевидно), а о том, что смерть - адаптивное ( важное для приспособленности вида) явление, которое достигается включением ( дерепрессией) или выключением ( репрессией) группы генов, несущих информацию о программе саморазрушения. [12]
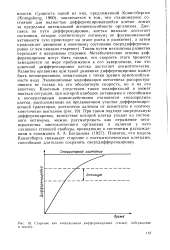 |
Старение как запредельная дифференцировка ( схема ( обсуждение в тексте. [13] |
Сущность одной из них, предложенной Кенигсбергом ( Konigsberg, 1960), заключается в том, что стационарное состояние для полностью дифференцировавшейся клетки лежит за пределами оптимальной жизнеспособности организма. Таким путем механизмы развития переходят в механизмы старения. Метаболические основы дифференцировки могут быть такими, что скорость этого процесса замедляется по мере приближения к его завершению, так что конечной дифференцировки клетка достигает асимптотически. Развитие организма при такой динамике дифференцировки может быть несовершенным, невыгодным с точки зрения приспособленности вида. Эволюционные модификации онтогенеза распространяются не только на его абсолютную скорость, но и на его кинетику. Понятно, что модель Конигсберга связывает старение с постмитотическими клетками, способными длительно сохранять сверхдифференцировку. [14]
Страницы: 1