Протеинкиназ
Cтраница 2
В 1968 г. Эдвин Кребс и его коллеги показали, что сАМР - за-виси мая протеинкиназа участвует в стимуляции глпкогенолиза. Связывание сАМР с регуляторными субъединицами вызывает диссоциацию их каталитических субъединиц, и последние, таким образом, активируются. Протеинкиназа, найденная в основном в нервных тканях, так называемая киназа типа II, фактически фосфорилирует свои собственные регуляторные субъединицы; такое автофосфорилирование не наблюдается у протеинкиназы типа I. Однако механизмы их активации одинаковы. [16]
Такие протеинкиназы содержат наряду с каталитическими субъединицами регуляторные субъединицы, содержащие центры узнавания цАМФ, который играет роль аллостерического активатора протеинкиназы. В результате этого возникает еще одна регуляторная ступень, предшествующая активации фосфорилазы. Принципиально отличаясь по своему химическому содержанию, эта ступень тем не менее работает по сходной схеме. Одновременно в клетке непрерывно функционирует фермент, катализирующий гидролиз цАМФ до АМФ - циклонуклеотид фосфогидролаза. Аденилатци-клаза вмонтирована в клеточную мембрану и. В частности, таким сигналом может служить гормон адреналин, активная наработка которого надпочечниками начинается в стрессовых ситуациях, когда организму требуется резкое усиление производства АТФ в связи с интенсификацией нервной деятельности, а зачастую и необходимостью совершить оперативно определенную механическую работу. [17]
Увеличение содержания егГМФ в печени через 30 мин, 6 - 9 и 24 ч после действия радиации может внести свой вклад в модификацию ядерных белков, поскольку известно, что цГМФ - зависимые протеинкиназы участвуют в фос-форилировании гистонов Н2А, Н2В, НЗ и Н4 [ Hashimoto E. [18]
Циклические нуклеотиды - 3 5 - АМФ ( цАМФ) и 3 5 - ГМФ ( цГМФ), образуемые соответственно в аденилатциклазной и гуанилатциклазной реакциях. Они активируют соответствующие протеинкиназы. Внутриклеточные эффекты этих циклических нук-леотидов часто противоположны по направленности. [19]
Данные о механизме действия АКТГ на синтез стероидных гормонов свидетельствуют о существенной роли аденилатциклазной системы. Последний активирует протеинкиназу, которая в свою очередь с участием АТФ осуществляет фосфорилирование холинэстеразы, превращающей эфиры холестерина в свободный холестерин, который поступает в митохондрии надпочечников, где содержатся все ферменты, катализирующие превращение холестерина в кортикостероиды. [20]
Эти Р.б. - протеинкиназы ( регулируют активность разл. [21]
Тжипаназолы А1 ( 16 - 7) и А2 ( 16 - 8) проявляют избирательную активность к различным видам патогенных грибов. Все тжипаназолы ингибируют протеинкиназу С, выделенную из мозга крыс. Обращает на себя внимание тот факт, что тжипаназолы ( 16 - 4) - ( 16 - 12) относятся к довольно редкому типу N-гликозилиро-ванных карбазолов. [22]
Образуются два очень важных для активации клеток компонента. Другой - диацилглице-рин, воздействует на протеинкиназу С. Таким образом происходит как бы раздвоение сигнала. [23]
Установлено, что эта киназа может существовать как в активной, так и в неактивной форме. Неактивная киназа фосфорилазы превращается в активную под влиянием фермента протеинкиназы ( киназа киназы фосфорилазы), и не просто протеинкиназы, а цАМФ - зависимой протеинкиназы. [24]
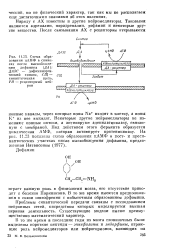 |
Схема образования цАМФ в синапсах после высвобояаде-ния дофамипа ( ДА. ДАЭС - дофаминэрги-ческий синапс, СЩ - синаптпческая щель, РН - рецепторньш нейрон. [25] |
Некоторые другие нейромедиаторы не изменяют ионные потоки, а активируют аденилатциклазу, связанную с мембраной. Под действием этого фермента образуется циклическая АМФ, которая активирует протеинкиназу. [26]
Таким образом, представленные данные о вторичных мессенджерах свидетельствуют о том, что каждой из этих систем посредников гормонального эффекта соответствует определенный класс протеинкиназ, хотя нельзя исключить возможности существования тесной связи между этими системами. Активность протеинкиназ типа А регулируется цАМФ, протеин-киназы G - цГМФ; Са2 - кальмодулинзависимые протеинкиназы находятся под контролем внутриклеточной [ Са2 ], а протеинкиназа типа С регулируется диацилглицеролом в синергизме со свободным Са2 и кислыми фосфолипидами. Повышение уровня какого-либо вторичного мес-сенджера приводит к активации соответствующего класса протеинкиназ и последующему фосфорилированию их белковых субстратов. В результате меняется не только активность, но и регуляторные и каталитические свойства многих ферментных систем клетки: ионных каналов, внутриклеточных структурных элементов и генетического аппарата. [27]
Установлено, что эта киназа может существовать как в активной, так и в неактивной форме. Неактивная киназа фосфорилазы превращается в активную под влиянием фермента протеинкиназы ( киназа киназы фосфорилазы), и не просто протеинкиназы, а цАМФ - зависимой протеинкиназы. [28]
ПТГ действует на клетки-мишени по мемб-рано-опосредованному механизму, причем это действие реализуется в почках, костной ткани и кишечнике. В клетках почечных канальцев, богатых рецепторами к ПТГ, происходит активация аденилатциклазы, а также синтез цАМФ, который активирует протеинкиназу и участвует в регуляции транспорта ионов Na, K и Са2 через клеточные мембраны. ПТГ оказывает множественное действие на костную ткань. Он опосредованно активирует ферменты коллаге-назу и глюкуронидазу, что вызывает деструкцию органических компонентов кости, в частности коллагена и гликозамингликанов. [29]
 |
Физиологические эффекты норад-ренапина и адреналина.| Структурные формулы норадреналина и адреналина. [30] |
Страницы: 1 2 3