Зацепление - цепь
Cтраница 2
Если молекулярный вес линейного полимера достаточно высок для появления зацепления цепей, то эффективный коэффициент трения для наибольших времен релаксации и вязкости установившегося течения равен Q o, в то время как для малых времен релаксации он равен просто 0 ( см. гл. Для полиизо-бутилена [18], так же как для концентрированных растворов полистирола [19] и поливинилацетата [20], Qr, по-видимому, не зависит от температуры, поскольку выражение (11.12) справедливо с довольно высокой точностью. С другой стороны, для некоторых полимеров метакрилата [21] Q, очевидно, уменьшается с повышением температуры, так что о /, применимое в конечной зоне, сильнее зависит от температуры, чем ат, применимое в переходной зоне. [16]
Топологическая нерегулярность возникает из-за наличия концов полимерных цепей и различных зацеплений цепей, окруженных сравнительно упорядоченным веществом, в котором присутствует ближний порядок. Морфологическая неоднородность вызвана существованием относительно больших зон, заметно выделяющихся по характеру расположения сегментов макромолекул, образующих эти зоны, и по их физическим свойствам. В кристаллизующихся полимерах эта неоднородность ведет к образованию кристаллических и аморфных областей, фибрилл и сферолитов. [17]
На заключительной стадии репликации кольцевых молекул часто остается одно или несколько зацеплений цепей исходной молеку-лы друг за друга. ДНК-гираза может расцепить зацепленные кольца, используя свою способность вносить временный двуцепочечный разрыв. Такая активность гиразы действительно существенна для репликации ДНК, поскольку в мутантах по гиразе на непермиссив-ной температуре наблюдается нерасхождение дочерних молекул кольцевых ДНК после репликации. Важно отметить, что топоизомеразы необходимы для завершения репликации не только кольцевых молекул, но и очень длинных линейных эукариотических хромосом: две очень длинные дочерние молекулы не могут разойтись достаточно быстро, поскольку после репликации оказываются запутанными подобно катенанам, образующимся на заключительной стадии репликации кольцевых ДНК. Действительно, мутанты эука-риот ( дрожжей) с нарушенной топоизомеразой II дефектны по расхождению дочерних хромосом в митозе. [18]
На заключительной стадии репликации кольцевых молекул часто остается одно или несколько зацеплений цепей исходной молекулы друг за друга. ДНК-гираза может расцепить зацепленные кольца, используя свою способность вносить временный двуцепочечный разрыв. Такая активность гиразы действительно существенна для репликации ДНК, поскольку в мутантах по гиразе на непермиссив-ной температуре наблюдается нерасхождение дочерних молекул кольцевых ДНК после репликации. Важно отметить, что топоизомеразы необходимы для завершения репликации не только кольцевых молекул, но и очень длинных линейных эукариотических хромосом: две очень длинные дочерние молекулы не могут разойтись достаточно быстро, поскольку после репликации оказываются запутанными подобно катенанам, образующимся на заключительной стадии репликации кольцевых ДНК. Действительно, мутанты эука-риот ( дрожжей) с нарушенной топоизомеразой II дефектны по расхождению дочерних хромосом в митозе. [19]
 |
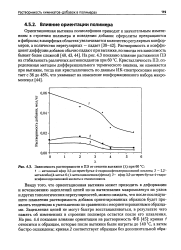
Характер изменения нагрузок в гусеничном ходовом механизме при установившемся движении и внедрении заборно-погрузочного органа. [20] |
Характер изменения усилий свидетельствует о значительной доле динамических составляющих, вызванных зацеплением цепи с ведущей звездочкой. [21]
Силы Рц не оказывают воздействия на зубья звездочек и не влияют на зацепление цепи со звездочкой. [22]
 |
Зависимость растворимости в ПЭ от степени вытяжки ( X при 60 С. [23] |
Ввиду того, что ориентационная вытяжка может приводить к деформации и исчезновению зацеплений цепей из-за вытягивания макромолекул из узлов и других топологических нерегулярностей, можно ожидать, что после последующего плавления растворимость добавок ориентированных образцов будет проявлять тенденцию к уменьшению по сравнению с неориентированными образцами. Зацепления цепей не могут быстро восстанавливаться, в результате чего память об изменениях в строении полимера остается после его плавления. [24]
 |
Структурная схема межцентровых расстояний с расположением звездочек внутри цепного контура.| Структурная схема межцентровых расстояний с расположением звездочек внутри и снаружи цепного контура. [25] |
При этом независимо от изменения конфигурации многоугольника и углов пересечения центры элементов зацепления цепи будут всегда оставаться в точках касания шаговых линий с делительными окружностями звездочек. [26]
Реакция окисления прежде всего вовлекает области полимера с меньшей плотностью упаковки, содержащей зацепления цепей, и может вести к исчезновению некоторых из них. [27]
Особенности формы зубьев и конструкции звездочек цепных передач определяются особенностями контакта между элементом зацепления цепи и зубом звездочки. Контактирующая с зубом часть элемента зацепления приводных пластинчатых втулочно-роликовых, грузовых пластинчатых, тяговых пластинчатых и тяговых разборных цепей имеет цилиндрическую форму, которая обеспечивает однородность условий зацепления звездочек и таких цепей, а значит и единство требований к форме зубьев и конструкции звездочек. [28]
Чрезмерное смещение звеньев от центра ухудшает условия работы и может привести к нарушению зацепления цепи со звездочкой. [29]
Допустимая величина относительного износа ( А / / /) ограничивается возможностью потери зацепления цепи со звездочкой, а также уменьшением прочности цепи. [30]
Страницы: 1 2 3 4