Открытая рамка - считывание
Cтраница 1
Открытая рамка считывания не содержит терминирующих кодонов и может транслироваться в белок. [1]
Механизм устойчивости ВКО к интерферону оставался неустановленным, пока не была обнаружена открытая рамка считывания K3L, кодирующая белок мол. Этот белок содержит аминокислотную последовательность, гомологичную N-концевой части эукариотиче-ского фактора инициации elF - 2a мол. N-концевые области обоих белков содержат 87 практически идентичных аминокислотных остатков, причем в положении 51 в обоих случаях находится серии, который в elF - 2a фосфорилируется активируемой интерфероном Р1 - киназой, что приводит к ингибированию синтеза белка в обработанных интерфероном клетках. КЗЬ-белок действует как конкурентный ингибитор фосфорилирования elF - 2a, обеспечивая устойчивость ВКО к интерферону, и если из генома ВКО удалить ген K3L или его часть, то вирус станет чувствительным к интерферону. Эта работа является важным этапом на пути создания более безопасных ВКО-векторов. [2]
Из трех возможных разбиений нуклеотидной последовательности на кодоны выбрать правильное часто удается по наличию при этом разбиении открытой рамки считывания - последовательности кодонов, среди которых на большом протяжении не встречается ко донов-терминаторов. [3]
Синтез VP1 при трансляции 16S мРНК начинается на одном из последующих AUG-триплетов; при этом используется другая, еще более протяженная открытая рамка считывания. [4]
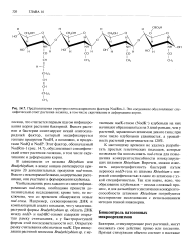 |
Предполагаемая структура олигосахаридного фактора NodRm-1. Это соединение обусловливает специфический ответ растения-хозяина, в том числе скручивание и деформацию корня. [5] |
Чтобы выяснить роль каждого из идентифицированных иоа - генов, необходимо провести дополнительные исследования; кроме того, не исключено, что со временем обнаружатся новые nod - гены. ДНК между nodD - и иоа йС - генами содержит открытую рамку считывания, а у быстрорастущей формы этой последовательности нет. [6]
РНК для Т - антигена и - антигена - эта разные молекулы, но образуются они из одного и того же класса ранних транскриптов. Различие имеется лишь на стыке этих двух районов: при образовании мРНК для - антигена в результате сплайсинга из первичного транскрипта удаляется 66 нуклеотидов, а при образовании мРНК для Т - антигена удаляется 346 нуклеотидов. В обеих мРНК инициаторный кодон предшествует участку сплайсинга, поэтому N-концевые последовательности Т - и - антиге-нов идентичны. В мРНК для - антигена терминирующий кодон также предшествует участку сплайсинга; имеющаяся открытая рамка считывания разрешает трансляцию относительно короткого полипептида. При образовании мРНК для Т - антигена этот терминирующий кодон удаляется в результате сплайсинга; возникает более протяженная открытая рамка считывания, и в состав Т - антигена входит уже примерно в четыре раза больше аминокислотных остатков. [7]
РНК для Т - антигена и / - антигена - это разные молекулы, но образуются они из одного и того же класса ранних транскриптов. Различие имеется лишь на стыке этих двух районов: при образовании мРНК для / - антигена в результате сплайсинга из первичного транскрипта удаляется 66 нуклеотидов, а при образовании мРНК для Т - антигена удаляется 346 нуклеотидов. В обеих мРНК инициаторный кодон предшествует участку сплайсинга, поэтому N-концевые последовательности Т - и / - антигенов идентичны. В мРНК для / - антигена терминирующий кодон также предшествует участку сплайсинга; имеющаяся открытая рамка считывания разрешает трансляцию относительно короткого полипептида. При образовании мРНК для Т - антигена этот терминирующий кодон удаляется в результате сплайсинга; возникает более протяженная открытая рамка считывания, и в состав Т - антигена входит уже примерно в четыре раза больше аминокислотных остатков. [8]
РНК для Т - антигена и - антигена - эта разные молекулы, но образуются они из одного и того же класса ранних транскриптов. Различие имеется лишь на стыке этих двух районов: при образовании мРНК для - антигена в результате сплайсинга из первичного транскрипта удаляется 66 нуклеотидов, а при образовании мРНК для Т - антигена удаляется 346 нуклеотидов. В обеих мРНК инициаторный кодон предшествует участку сплайсинга, поэтому N-концевые последовательности Т - и - антиге-нов идентичны. В мРНК для - антигена терминирующий кодон также предшествует участку сплайсинга; имеющаяся открытая рамка считывания разрешает трансляцию относительно короткого полипептида. При образовании мРНК для Т - антигена этот терминирующий кодон удаляется в результате сплайсинга; возникает более протяженная открытая рамка считывания, и в состав Т - антигена входит уже примерно в четыре раза больше аминокислотных остатков. [9]
РНК для Т - антигена и / - антигена - это разные молекулы, но образуются они из одного и того же класса ранних транскриптов. Различие имеется лишь на стыке этих двух районов: при образовании мРНК для / - антигена в результате сплайсинга из первичного транскрипта удаляется 66 нуклеотидов, а при образовании мРНК для Т - антигена удаляется 346 нуклеотидов. В обеих мРНК инициаторный кодон предшествует участку сплайсинга, поэтому N-концевые последовательности Т - и / - антигенов идентичны. В мРНК для / - антигена терминирующий кодон также предшествует участку сплайсинга; имеющаяся открытая рамка считывания разрешает трансляцию относительно короткого полипептида. При образовании мРНК для Т - антигена этот терминирующий кодон удаляется в результате сплайсинга; возникает более протяженная открытая рамка считывания, и в состав Т - антигена входит уже примерно в четыре раза больше аминокислотных остатков. [10]
Известны и другие вирусы как с непрерывным, так и сегментированным ( -) РНК-геномом. И хотя какие-то детали механизмов их репликации / транскрипции могут отличаться от только что описанных, общность основных принципов прослеживается удивительно хорошо. Но на одной группе вирусов следует остановиться особо. Речь идет о флебовирусах, которые относят к обширному семейству буньявирусов. Все буньявирусы имеют геном, состоящий из трех сегментов. У обычных буньявирусов, подобно вирусу гриппа, все эти сегменты представляют собой молекулы ( -) РНК, на каждой из которых как на матрице синтезируются как субгеномные мРНК с гетерогенными кэпированными 5 -концами, происходящими из клеточных РНК, так и полноразмерные молекулы () РНК - Здесь рассмотрим в первую очередь малый ( S) сегмент буньявирусов; у обычных представителей этого семейства он содержит информацию о двух белках, которые считываются с одной молекулы субгеномной мРНК благодаря использованию разных инициатор-ных триплетов и разных открытых рамок считывания. [11]
Известны и другие вирусы как с непрерывным, так и сегментированным ( -) РНК-геномом. И хотя какие-то детали механизмов их репликации / транскрипции могут отличаться от только что описанных, общность основных принципов прослеживается удивительно хорошо. Но на одной группе вирусов следует остановиться особо. Речь идет о флебовирусах, которые относят к обширному семейству буньявирусов. Все буньявирусы имеют геном, состоящий из трех сегментов. У обычных буньявирусов, подобно вирусу гриппа, все эти сегменты представляют собой молекулы ( -) РНК, на каждой из которых как на матрице синтезируются как субгеномные мРНК с гетерогенными кэпированными 5 -концами, происходящими из клеточных РНК, так и полноразмерные молекулы () РНК. Здесь рассмотрим в первую очередь малый ( S) сегмент буньявирусов; у обычных представителей этого семейства он содержит информацию о двух белках, которые считываются с одной молекулы субгеномной мРНК благодаря использованию разных инициатор-ных триплетов и разных открытых рамок считывания. [12]
Страницы: 1