Расплетение
Cтраница 3
В синтезируемой молекуле РНК отдельные мононуклеотиды, как и в ДНК, связаны между собой 3 - 5 -фосфодиэфирными мостиками. Кроме того, сам механизм действия фермента РНК-полимеразы во многом совпадает с таковыми ДНК-полимеразы: синтез также идет в направлении 5 - 3, цепь РНК имеет полярность, противоположную цепи предобразованной ДНК. Однако выявлены и существенные различия. ДНК; в опытах in vitro обе цепи ДНК копируются РНК-полимеразой; in vivo транскрибируется, вероятнее всего, только одна цепь ДНК. Предполагают, что РНК-полимераза связывается с одной цепью нативной ДНК в определенной точке, вызывая расплетение биспиральной структуры на ограниченном участке, где и происходит синтез РНК. [31]
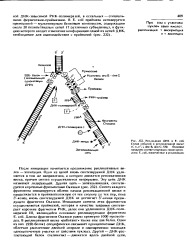 |
Репликация ДНК в Е. coli. Схема событий в репликотиннон вилке. п, п, п, i, dna В, dna С, SSb белковые продукты соответствующих генов хромо сомы Е. coli. вовлеченные в репликацию. [32] |
После инициации начинается продвижение репликативных вилок - элонгация. Одна из цепей вновь синтезируемой ДНК удлиняется в том же направлении, а котором движется репликативная вилка, причем синтез осуществляется непрерывно. Эту цепь ДНК называют лидирующей. Синтез каждого фрагмента инициируется вблизи начала репликационной вилки и продолжается в противоположную от нее сторону до тех пор, пока З - конец вновь синтезируемой ДНК не достигнет 5 -конца предыдущего фрагмента Оказаки. Длины фрагментов Оказаки равны примерно 1000 нуклеоти-доа. В репликативной вилке работают также еще два белка. Один из них ( SSb белок) специфически связывает одноцепочечные ДНК, облегчая расплетение двойной спирали и одновременно защищая одно цепочечные участки от действия нуклеаз. Другой - ДНК-рас-плетающий белок ( хеликазы) - движется вдоль двойной цепи. [33]
Варшавский и Евдокимов изучали расплетание ДНК методом теплового удара ( ср. Кинети: ческие кривые свидетельствуют о наличии двух или даже трех стадий структурного перехода. В первой, быстрой, стадии возникает почти весь гиперхромный эффект. Полупериод всего перехода меняется в зависимости от условий в пределах от нескольких секунд до нескольких десятков секунд. Константа скорости следует уравнению Аррениуса. Расплетание спирали происходит с наибольшей скоростью при экстремальных рН - зависимость энергии активации от рН колоколообразна. Максимальное значение Е 170 ккал / моль отвечает рН 7 5 ( ионная сила 0 18 - 0 25); оно падает до 20 - 25 ккал / моль при рН 3 и 10 5, Авторыинтер-претируют первую ста дню как образование неподвижных петель и считают, что полное расплетение происходит во второй стадии. Время расплетания возрастает с увеличением молекулярного веса. [34]
Репликация всегда предшествует делению клетки и начинается с распле-тения двойной спирали ДНК. Это осуществляется при помощи ферментов хе-ликаз, которые перемещаются вдоль цепей ДНК и раскручивают их. Процесс расплетения спиралей ДНК является энергозависимым и требует затраты АТФ. Интенсивное раскручивание ДНК может привести к образованию дополнительных витков. Это явление носит название положительной сверхспира-лизации или сверхскрученности и устраняется при помощи ферментов топоизо-мераз. В частности, топоизомераза II осуществляет релаксацию положительной сверхспирализации за счет образования отрицательных сверхвитков. Топоизомераза II также носит название гираза. После расплетения двух нитей ДНК необходимо их стабилизировать в этом состоянии. Оказалось, что существует специальный белок, специфично связывающийся с одной из нитей ДНК и препятствующий обратной рекомбинации в двойную спираль. Таким образом, расплетение ДНК и образование репликативных вилок является достаточным основанием ( при наличии ферментов и субстратов репликации) для удвоения ДНК. [35]
Репликация всегда предшествует делению клетки и начинается с распле-тения двойной спирали ДНК. Это осуществляется при помощи ферментов хе-ликаз, которые перемещаются вдоль цепей ДНК и раскручивают их. Процесс расплетения спиралей ДНК является энергозависимым и требует затраты АТФ. Интенсивное раскручивание ДНК может привести к образованию дополнительных витков. Это явление носит название положительной сверхспира-лизации или сверхскрученности и устраняется при помощи ферментов топоизо-мераз. В частности, топоизомераза II осуществляет релаксацию положительной сверхспирализации за счет образования отрицательных сверхвитков. Топоизомераза II также носит название гираза. После расплетения двух нитей ДНК необходимо их стабилизировать в этом состоянии. Оказалось, что существует специальный белок, специфично связывающийся с одной из нитей ДНК и препятствующий обратной рекомбинации в двойную спираль. Таким образом, расплетение ДНК и образование репликативных вилок является достаточным основанием ( при наличии ферментов и субстратов репликации) для удвоения ДНК. [36]
Исключение составляют нек-рые вирусы, в составе к-рых обнаружены одноцепо-чечные ДНК, также линейные или кольцевые. В клетках прокариот ДНК организована в одну хромосому - нуклеоид - и представляет единую макромолекулу с мол. ДНК присутствуют в плазм идах. В клетках эука-риот ДНК находится гл. Полагают, что хромосома эукариот, подобно бактериальной, состоит из одной молекулы ДНК с очень высокой мол. Минимальное для данного вида кол-во ДНК содержат половые клетки, имеющие гаплоидный набор хромосом. ДНК, как правило, вдвое больше, чем соответствует диплоидному набору. Относит, содержание ДНК определяется видовыми особенностями и функциональным состоянием клетки, составляя обычно неск. Биосинтез ДНК осуществляется путем матричного синтеза ( в основе лежат закономерности образования комплементарных пар) по полу консервативному механизму. Репликация хромосомной ДНК в делящейся клетке начинается с локального расплетения двойной спирали и образования репликативной вилки, в чем принимают участие специфич. [37]
Страницы: 1 2 3