Реакция - трансэтерификация
Cтраница 1
Реакция трансэтерификации изучена довольно мало, но Мерорта [45 ] успешно использовал ее для получения третичных алкоксидов циркония и гафния, и она, по-видимому менее чувствительна к стерическим затруднениям по сравнению с реакцией спиртового обмена. Это может быть в том случае, если начальная стадия трансэтерификации заключается в координировании карбонильного кислорода сложного эфира, а не алкоксильного кислорода. [1]
Механизм этого процесса не ясен. Возможно, вставки интронов происходили на уровне РНК - Если процесс вырезания интрона с помощью реакций трансэтерификации термодинамически обратим ( см. гл. [2]
Механизм этого процесса не ясен. Возможно, вставки интронов происходили на уровне РНК. Если процесс вырезания интрона с помощью реакций трансэтерификации термодинамически обратим ( см. гл. [3]
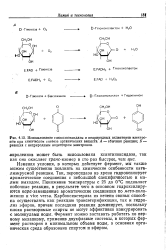 |
Использование глюкозооксидазы и неприродных акцепторов электро-ов при химическом синтезе органических веществ. А - обычная реакция. Б - реакция с неприродным акцептором электронов. [4] |
Изменяя условия, в которых действует фермент, мы также можем существенно повлиять на химические особенности катализируемой реакции. Так, пероксидаза из хрена гидроксилирует ароматические соединения с небольшой специфичностью и малым выходом. Понижение температуры с 25 до 0 С подавляет побочные реакции, в результате чего в основном гидроксилиру-ются / шра-замещенные ароматические соединения по мета-поло-жению и vice versa. Карбоксиэстераза из печени свиньи способна осуществлять как реакцию трансэтерификации, так и гидролиз эфиров, причем последняя реакция доминирует, поскольку плохо растворимые спирты не могут эффективно конкурировать; молекулами воды. Фермент можно заставить работать по первому механизму, применив двухфазные системы, в которых фермент растворен в минимальном объеме воды, а основная органическая среда образована спиртом и эфиром. [5]
Ферментативная машина сплайсинга, локализованная в ядре, изучена недостаточно, однако ряд важных результатов получен при исследовании превращений предшественников индивидуальных мРНК ( например, р-глобина) в ядерных экстрактах in vitro. Прежде всего происходит разрыв нуклеотидной последовательности на границе экзона 1 и интрона с образованием 5 -конца, представленного фос-форилированным гуанозином - каноническим нуклеотидом левого ( 5 -конца) интрона. Следующая ( или сопряженная с предыдущей) реакция заключается во взаимодействии 5 -конца удаляемой последовательности с 2 -гидроксилом аденозина интрона с образованием. Образование такой структуры можно также рассматривать как результат реакции трансэтерификации, в результате которой 2 - ОН-группа аденозина атакует фосфодиэфирную связь, замещая З - ОН-группу экзона 1 Теперь экзон 1 представлен отдельной молекулой РНК, удерживаемой вместе с другой молекулой, содержащей лассд-интрон и экзон 2, в едином комплексе с белками, участвующими в реакции. По-видимому, образование лассо является условием для завязывания ковалентной связи З - ОН-группы линейной молекулы экзона 1 с экзоном 2 в районе сайта сплайсинга; эта реакция также представляет собой трансэтерификацию, когда З - ОН-группа экзона 1 атакует фосфодиэфирную связь и замещает З - ОН-группу интрона; последовательность интрона выщепляется из РНК в форме лассо. В результате число фосфодиэфирных связей в составе субстрата и продукта не изменяется. [6]
Ферментативная машина сплайсинга, локализованная в ядре, изучена недостаточно, однако ряд важных результатов получен при исследовании превращений предшественников индивидуальных мРНК ( например, р-глобина) в ядерных экстрактах in vitro. Прежде всего происходит разрыв нуклеотидной последовательности на границе экзона 1 и интрона с образованием 5 -конца, представленного фосфор ил ированным гуанозином - каноническим нуклеотидом левого ( 5 -конца) интрона. Следующая ( или сопряженная с предыдущей) реакция заключается во взаимодействии 5 -конца удаляемой последовательности с 2 -гидроксилом аденозина интрона с образованием; 2 - 5 -фосфодиэфирной связи. Образование такой структуры можно также рассматривать как результат реакции трансэтерификации, в результате которой 2 - ОН-группа аденозина атакует фосфодиэфирную связь, замещая З - ОН-группу экзона 1 Теперь экзон 1 представлен отдельной молекулой РНК, удерживаемой вместе с другой молекулой, содержащей лассб-интрон и экзон 2, в едином комплексе с белками, участвующими в реакции. По-видимому, образование лассо является условием для завязывания ковалентной связи З - ОН-группы линейной молекулы экзона 1 с экзоном 2 в районе сайта сплайсинга; эта реакция также представляет собой трансэтерификацию, когда З - ОН-группа экзона 1 атакует фосфодиэфирную связь и замещает З - ОН-группу интрона; последовательность интрона выщепляется из РНК в форме лассо. В результате число фосфодиэфирных связей в составе субстрата и продукта не изменяется. [7]
Ферментативная машина сплайсинга, локализованная в ядре, изучена недостаточно, однако ряд важных результатов получен при исследовании превращений предшественников индивидуальных мРНК ( например, р-глобина) в ядерных экстрактах in vitro. Прежде всего происходит разрыв нуклеотидной последовательности на границе экзона 1 и интрона с образованием 5 -конца, представленного фос-форилированным гуанозином - каноническим нуклеотидом левого ( 5 -конца) интрона. Следующая ( или сопряженная с предыдущей) реакция заключается во взаимодействии 5 -конца удаляемой последовательности с 2 -гидроксилом аденозина интрона с образованием; 2 - 5 -фосфодиэфирной связи. Образование такой структуры можно также рассматривать как результат реакции трансэтерификации, в результате которой 2 - ОН-группа аденозина атакует фосфодиэфирную связь, замещая З - ОН-группу экзона 1 Теперь экзон 1 представлен отдельной молекулой РНК, удерживаемой вместе с другой молекулой, содержащей лассд-интрон и экзон 2, в едином комплексе с белками, участвующими в реакции. По-видимому, образование лассо является условием для завязывания ковалентной связи З - ОН-группы линейной молекулы экзона 1 с экзоном 2 в районе сайта сплайсинга; эта реакция также представляет собой трансэтерификацию, когда З - ОН-группа экзона 1 атакует фосфодиэфирную связь и замещает З - ОН-группу интрона; последовательность интрона выщепляется из РНК в форме лассо. В результате число фосфодиэфирных связей в составе субстрата и продукта не изменяется. [8]
Реакции происходят и между компонентами древесины и дистиллята. Обычно это реакции этерификации, но теоретически они могут включать также реакции окисления и ацетилирования. В ходе выдержки вследствие этерификации свободных кислот этиловым спиртом увеличивается концентрация эфиров. Считается также, что при выдержке могут происходить и реакции трансэтерификации, которые при наличии избытка этилового спирта способствуют образованию этиловых эфиров. [9]
Пожалуй, это единственные из известных макромолекул, которые наделены как информационной, так и каталитической функцией. Оказалось, что некоторые РНК осуществляют посттранскрипционный процессинг, катализируя самосплайсинг, т.е. участвуют в разрезании и удалении интронов. Наделенные рядом свойств истинных и эффективных катализаторов рибозимы участвуют в двух типах реакций: в гидролизе ( разрыве) фосфодиэфирной связи и в реакциях трансэтерификации. В качестве субстрата могут служить, помимо собственного, предшественник ( про - РНК) и другие молекулы РНК. Сейчас интенсивно изучается третичная структура рибозимов, а первичная и вторичная структуры ряда из них уже расшифрованы. Эти исследования, несомненно, интересные сами по себе, могут пролить свет и на пути развития биологической эволюции. [10]
Страницы: 1